By the classification of any series of objects, is meant the actual, or ideal, arrangement together of those which are like and the separation of those which are unlike; the purpose of this arrangement being to facilitate the operations of the mind in clearly conceiving and retaining in the memory, the characters of the objects in question.
Thus, there may be as many classifications of any series of natural, or of other, bodies, as they have properties or relations to one another, or to other things; or, again, as there are modes in which they may be regarded by the mind: so that, with respect to such classification as we are here concerned with, it might be more proper to speak of a classification than of the classification of the animal kingdom.
The preparations in the galleries of the Museum of the Royal College of Surgeons are arranged upon the basis laid down by John Hunter, whose original collection was intended to illustrate the modifications which the great physiological apparatuses undergo in the [2] animal series: the classification which he adopted is a classification by organs, and, as such, it is admirably adapted to the needs of the comparative physiologist.
But the student of the geographical distribution of animals, regarding animated creatures, not as diverse modifications of the great physiological mechanism, but in relation to one another, to plants and to telluric conditions, would, with equal propriety, dispose of the contents of a Zoological Museum in a totally different manner; basing his classification, not upon organs, but on distributional assemblages. And the pure palæontologist, looking at life from yet another distinct point of view, would associate animal remains together on neither of these principles, but would group them according to the order of their succession in Time.
Again, that classification which I propose to discuss in the present pages, is different from all of these: it is meant to subserve the comprehension and recollection of the facts of animal structure; and, as such, it is based upon purely structural considerations, and may be designated a Morphological Classification. I shall have to consider animals, not as physiological apparatuses merely; not as related to other forms of life and to climatal conditions; not as successive tenants of the earth; but as fabrics, each of which is built upon a certain plan.
It is possible and conceivable that every animal should have been constructed upon a plan of its own, having no resemblance whatsoever to the plan of any other animal. For any reason we can discover to the contrary, that combination of natural forces which we term Life might have resulted from, or been manifested by, a series of infinitely diverse structures: nor, indeed, would anything in the nature of the case lead us to suspect a community of organization between animals, so different in habit and in appearance, as a porpoise and a gazelle, an eagle and a crocodile, or a butterfly and a lobster. Had animals been thus independently organized, each working out its life by a mechanism peculiar to itself, such a classification as that which is now under contemplation would obviously be impossible; a morphological, or structural, classification plainly implying morphological or structural resemblances in the things classified.
[3] As a matter of fact, however, no such mutual independence of animal forms exists in nature. On the contrary, the different members of the animal kingdom, from the highest to the lowest, are marvellously interconnected. Every animal has a something in common with all its fellows: much, with many of them; more, with a few; and, usually, so much with several, that it differs but little from them.
Now, a morphological classification is a statement of these gradations of likeness which are observable in animal structures, and its objects and uses are manifold. In the first place, it strives to throw our knowledge of the facts which underlie, and are the cause of, the similarities discerned into the fewest possible general propositions–subordinated to one another, according to their greater or less degree of generality; and in this way it answers the purpose of a memoria technica, without which the mind would be incompetent to grasp and retain the multifarious details of anatomical science.
But there is a second and even more important aspect of morphological classification. Every group in that classification is such in virtue of certain structural characters, which are not only common to the members of the group, but distinguish it from all others; and the statement of these constitutes the definition of the group.
For example: if a fragmentary fossil be discovered, consisting of no more than a ramus of a mandible and that part of the skull with which it articulated, a knowledge of this law may enable the palæontologist to affirm, with great confidence, that the animal of which it formed a part suckled its young and had non-nucleated red blood-corpuscles; and to predict that, should the back part of that skull be discovered, it will exhibit two occipital condyles and a well-ossified basioccipital bone.
Deductions of this kind, such as that made by Cuvier in the famous case of the fossil opossum of Montmartre, have often been verified, and are well calculated to impress the vulgar imagination; so that they have taken rank as the triumphs of the anatomist. But it should carefully be borne in mind, that, like all merely empirical laws, which rest upon a comparatively narrow observational basis, the reasoning from them may at any time break down. If Cuvier, for example, had had to do with a fossil Thylacinus instead of a fossil Opossum, he would not have found the marsupial bones, though the inflected angle of the jaw would have been obvious enough. And so, though, practically, any one who met with a characteristically mammalian jaw would be justified in expecting to find the characteristically mammalian occiput associated with it; yet, he would be a bold man indeed, who should strictly assert the belief which is implied in this expectation, viz., that at no period of the world's history did animals exist which combined a mammalian occiput with a reptilian jaw, or vice versa.
Not that it is to be supposed that the correlations of structure expressed by these empirical laws are in any sense accidental, or other than links in the general chain of causes and effects. Doubtless there is some very good reason why the characteristic occiput of a Mammal should be found in association with mammæ and non-nucleated blood-corpuscles; but it is one thing to admit the causal connection of these phenomena with one another, or with some third; and another thing to affirm that we have any knowledge of that causal connexion, or that [5] physiological science, in its present state, furnishes us with any means of reasoning from the one to the other.
Cuvier, the more servile of whose imitators are fond of citing his mistaken doctrines as to the nature of the methods of palæontology against the conclusions of logic and of common sense, has put this so strongly that I cannot refrain from quoting his words.1
Morphological classification, then, acquires its highest importance as a statement of the empirical laws of the correlation of structures; and its value is in proportion to the precision and the comprehensiveness with which those laws, the definitions of the groups adopted in the classification, are stated. So that, in attempting to arrive at clear notions concerning classification, the first point is to ascertain whether any, and if so, what groups of animals can be established, the members of which shall be at once united together and separated from those of all other groups, by well-defined structural characters. And it will be most convenient to commence the inquiry with groups of that order which are commonly called Classes, and which are enume[3]rated in an order and arrangement, the purpose of which will appear more fully by and by, in the following table.
TABLE OF THE CLASSES OF THE ANIMAL KINGDOM.
The Limits of the Four Cuvierian Sub-Kingdoms are indicated by the Brackets and Dotted Line.
Gregarinda.
Rhizopoda (?).
Spongida.
Infusoria. |
} | Radiata. |
|
Hydrozoa
Actinozoa.
Polyzoa.
Brachipoda.
Ascidionida.
Lamellibranchiata.
Branchiogasterpoda.
Pulmogasteropoda.
Pteropoda.
Cephalopoda. |
} | Mollusca. |
|
Echionodermata.
Scoleceida (?).
Annelida.
Crustacea.
Arachnida.
Myriapoda.
Insecta. |
} | Articulata. |
|
Pisces.
Amphibia.
Reptilia.
Aves.
Mammalia. |
} | Vertebrata. |
[...]
1 'Ossemens fossiles,' ed. 4me, tome 1', p. 184.
[87]
Lecture VI. On the Classification of Animals.
The Subdivisions of the Mammalia Larger than Orders.
In my last lecture I endeavoured to point out the grounds upon which naturalists have arrived at the conclusion that the classes of the Animal Kingdom may be arranged together in larger groups or divisions, such as have been termed "provinces" and "sub-kingdoms." If the time at my disposal for the consideration of Classification permitted me to do so, I should now, in the logical order of my discourse, take the opposite course; and turning again to the list of classes, I should endeavour to indicate in what manner they must be subdivided into sub-classes, orders, and lesser divisions. But it is needless to say that such a task as this would require many lectures, while I have only one to dispose of; and I propose to devote that one to a consideration of the classification of that class, which is in many respects the most interesting and the most important of any in the Animal Kingdom,–the class Mammalia.
A great many systems of classification of the Mammalia have been proposed, but, as any one may imagine from the nature of the case, only those which have been published within the last forty or fifty years, or since our knowledge of the anatomy of these animals has approached completeness, have now any scientific standing-ground. I do not propose to go into the history of those older systems, which laboured more or less under the disqualification of being based upon imperfect know[88]ledge, but I shall direct your attention at once to that important step towards dividing the Mammalia into large groups, which was taken by the eminent French anatomist, M. de Blainville, so far back as the year 1816;. M. de Blainville pointed out that the Mammalia might be divided into three primary groups, according to the character of their reproductive organs, especially the reproductive organs of the female. He divided them into "Ornithodelphes," "Didelphes," "Monodelphes;" or, as we might term them, Ornithodelphia, Didelphia, Monodelphia. Now, I do not mean to assert that M. de Blainville defined these different groups in a manner altogether satisfactory, or strictly in accordance with all the subsequently discovered facts of science, but his great knowledge and acute intuition led him to perceive that the groups thus named were truly natural divisions of the Mammalia. And the enlargement of our knowledge by subsequent investigation seems to me, in the main, only to have confirmed De Blainville's views.
The division of the Ornithodelphia comprises those two remarkable genera of Mammals, as isolated in geographical distribution as in structure, –Ornithorhynchus and Echidna,– which constitute the order Monotremata.
In these animals the angle of the lower jaw is not inflected, and the jaws are devoid of true teeth, one of the two genera only (Ornithorhynchus) possessing horny plates in the place of teeth. The coracoid bone extends from the scapula to the sternum, with which it is articulated, as in birds and most reptiles, and, as in many of the latter, there is an episternal bone. There is no marsupial pouch, though bones wrongly termed "marsupial" are connected with the pelvis. But it is to the structure of the female reproductive organs that the Ornithodelphia owe their name. The oviducts, enlarged below into uterine pouches, but opening separately from one another, as in oviparous vertebrates, debouch, not into a distinct vagina, but into a cloacal chamber, common to the urinary and genital products and to the fæces. The testes of the male are abdominal in position throughout life, and the vasa deferentia open into the cloaca, and not into a distinct urethral passage. The penis is indeed traversed by an [89] urethral canal, but it is open and interrupted at the root of that organ. In both sexes, the ureters pour the renal secretion, not into the bladder, which is connected with the upper extremity of the cloaca, but into the latter cavity itself.
In the brain, the corpus callosum is inconspicuous, though the question how far it can properly be said to be absent requires much more thorough investigation than it has yet received.1 We are but very imperfectly acquainted with the reproductive processes of these animals, but it is asserted that the young are devoid of a placenta. The mammary gland has no nipple.
Like the Ornithodelphia, the division Didelphia contains but a single order, the Marsupialia, the great majority of which, like the Ornithodelphia, inhabit Australia. They almost all have the angle of the lower jaw inflected, and all possess true teeth. The coracoid is, as in the higher Mammals, anchylosed with the scapula, and is not articulated with the sternum. All have the so-called "marsupial" bones or cartilages–ossifications, or chondrifications, of the internal tendon of the external oblique muscle of the abdomen–and the females of almost all possess a fold of the skin of the abdomen above the pubis, constituting a "marsupium," or pouch, within which the young are nourished and protected in their early, helpless condition.
The oviducts open into vaginæ, which are more or less completely divided into two separate passages. The testes of the [90] males are lodged in a scrotum, which is suspended in front of the penis; and the vasa deferentia open into a complete and continuous urethra, which is also the passage by which the urine escapes from the bladder, and is perfectly distinct from the passage for the fæces, though the anus and the termination of the urethro-sexual canal are embraced by the same sphincter.
The corpus callosum is comparatively small, as in the Ornithodelphia.
It is stated that the allantois of the embryo is arrested in its development, and gives rise to no placenta. The umbilical sac is said to acquire a large proportional size; but whether it plays the part of a placenta for the short period of intra-uterine life, or not, is unknown.
The young are born of very small size, and in a singularly imperfect condition; but being transferred to the marsupium, and becoming attached to a long nipple, they are supplied with milk until they are able to provide for themselves–the milk being, at first, forced into their mouths by the action of a muscle spread over the mammary gland.
In the Monodelphia, the angle of the lower jaw is not inflected, and they may or may not be provided with teeth. They never possess "marsupial" bones. The uterine dilatation of the oviducts is always considerable, and whether they have common or distinct apertures, the vagina is a single tube, though it may be partially divided by a septum. The testes may vary much in position; but, if they are lodged in a scrotal pouch, it is never pendulous by a narrow neck in front of the penis, as in the Didelphia.
The urinary bladder opens into a distinct urethra, which, directly or indirectly, receives the vasa deferentia in the male.
The corpus callosum is very variable in its development, commonly attaining a much larger size than in the preceding groups; the optic lobes are divided into four portions.
The young are nourished within the uterus until such time as they are competent to suck milk from the teats of the parent, to which end the chorion always develops processes or villi, which are well supplied with vessels brought to them by the allantois. These processes becoming interlaced more or less [91] closely with corresponding vascular developments of wall of the uterus (and so forming a "placenta"), an interchange of constituents takes place between the fœtal and the maternal blood, through the separating walls of the fœtal and maternal vessels. In this manner, throughout its prolonged intra-uterine life, the Monodelphian is supplied with nourishment and gets rid of its effete products.
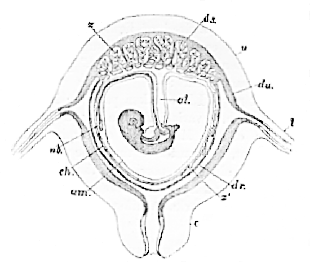
Fig. 39.–Diagrammatic section of a human pregnant uterus, with the contained ovum (Longet).
u, uterus; l, oviduct; c, cervix uteri; du, decidua uteri; dr, decidua reflexa; ds, decidua seretina; ch, chorion; am, amnion; al, allantois; ub, umbilical vesicle; x, villi which form the fœtal part of the placenta; x 1, villi over the rest of the chorion, which take no part in the placental function in man.
As the three groups instituted by De Blainville are capable of being thus clearly differentiated one from the other, the distinctions between them having been only more and more clearly brought out by the subsequent progress of knowledge, I can see no ground for refusing to adopt his classification, or for denying him that credit to which he is fairly entitled for apprehending these distinctions. Certainly, the later proposition, to divide Mammals into two great groups only, Placentalia and Implacentalia, cannot be regarded as any improvement upon De Blainville's system, as it ignores the important fact that the [92] two divisions of the Implacentalia are separated by characters of fully as great importance as those which distinguish the Placentalia and Implacentalia.
But whether the Ornithodelphia and the Didelphia are regarded (as I believe they ought to be) as two of the three primary separate "sub-classes" of the class Mammalia, or whether they are looked upon only as subdivisions of the Implacentalia, there is no doubt that they are, and will remain, distinct natural assemblages, the subdivisions of which present no very great difficulties.
It is otherwise with the sub-class Monodelphia–which contains at least a dozen orders, the arrangement of which into groups, not only in detail, but in principle, is, and long has been, a subject of much difficulty, and consequently of controversy
Sir Everard Home2 is commonly quoted as the originator of one of the two systems of classification in vogue at the present day; but his vague statements and confused notions respecting the varying characters of the placenta of the Monodelphia hardly entitle him to that honour, which, in my opinion, belongs rather to that eminent man, Karl Ernst von Baer, of whom it can be truly said that he has touched no subject without throwing a flood of light upon it. Towards the end of his famous essay, "Untersuchungen über die Gefäsverbindung zwischen Mutter und Frucht," published in 1828, the following passage occurs:–
"In the first place, I have taken pains to show that the ova of mammals are only variations of a single type; and if we except the ova of the Marsupials, concerning which I can form no judgment, all consist of the same parts; all have a placenta; and, in all, some portion of the chorion is smooth. The fœtal placenta consists everywhere of the same elements but offers the most remarkable differences in its external disposition. It is either–
1. Merely applied to the maternal placenta, and
(a) continuous and zone-like. First form.
(b) divided into many parts. Second form.
[93] Or 2. It and the maternal placenta grow together, and they lie,
(a) in a zone round the egg. Third form.
(b) at one end of it. Fourth form.
These differences, however, are developed gradually, and, at first, are less marked."
The first form, described in the text of the world is that met with in the pig. It is what is now commonly termed a diffused placenta; but Von Baer, more accurate than most of his successors, indicates the confinement of the placental villi to the middle of the chorion–its prolonged poles remaining bare–by the term "gürtelförmig," zone-like. The second form is that exemplified by the cow and sheep, the cotyledonary placenta. The third is the carnivorous placenta, termed zonular. The fourth is the placenta of man, called now-a-days discoidal.
The most important circumstance pointed out by Von Baer, however, is one which has been greatly overlooked, if not wholly ignored, in subsequent discussions–the fact that the differences in the form of mammalian placentæ are subsidiary if compared with their differences in structure, more particularly in regard to the extent to which a maternal element enters into their composition.
Eschricht, in the admirable memoir, "De Organis quæ Respirationi et Nutritioni Fœtus Mammalium inserviunt," which he. published in 1837, repeats the ideas of Von Baer, apparently without being aware of the fact, and enlarges upon them as follows (p. 30):–
"Restat, ut succinctam expositionem Mammalium afferamus secundum varias quæ in iis observantur, placentæ formas.
"A ceteris omnibus mammalibus Marsupialia et Monotremata separanda sunt, quibus nulla est placenta. Cætera omnia in duas familias dividenda, quarum alteri placenta uterine caduca, alteri non caduca est. Huie Mammalia primata et ungulata omnia adnumeranda sunt, inter quæ Ruminantia ob singularem cotyledorum formam cæteris opponi possunt.
"In mammalibus placentam uterinam caducam habentibus [94] tres mihi occurrere videntur placentæ typi, quorum primus gliribus, secundus feris, tertius simiis et homini proprius est."
In this passage Mammals are clearly divided, in the first place, into placental and implacental; and the former are then subdivided into those which have a non-caducous and those which have a caducous uterine placenta. The Cetacea and Ungulate Mammals constitute the former group; the Rodents, Carnivores, Apes, and Men the latter.
In 1843, an accomplished English zoologist, Mr. Waterhouse, published a highly instructive paper on the "Classification of the Mammalia,"3in which the following passage occurs:–
"Taking the general form of the Brain into consideration the placental Mammalia would appear divisible into two sections: first, those in which the cerebrum is generally of a rounded form, obtuse in front and provided with distinct convolutions; and secondly, those in which the cerebrum is destitute of convolutions, or nearly so, and usually contracted in front. The first division would contain the Quadrumana, Carnivora, Cetacea, Pachydermata, and Ruminantia, and the second would contain the Cheiroptera, Insectivora, Edentata, and Rodentia."
But although Mr. Waterhouse puts forward thus clearly the facts upon which a cerebral classification of the Mammalia might be based, he immediately afterwards, with his customary judgment, expresses great doubt as to the value of any such classification.
"But are we in a condition to take for a basis of classification of the Mammalia the structure of the brain? I think not, though, in the case of the Marsupalia, it has afforded characters serving to separate that from other sections, and to indicate its proper position in the system. I am not prepared to follow those naturalists who would, in the present state of information, take this organ as one of primary importance in the distribution of the orders of the placental series of Mammals. I cannot adopt the two great sections of this series as apparently [95] indicated by the smooth and anteriorly contracted cerebrum on the one hand, and the convoluted cerebrum, with its rounded anterior portion, on the other. Were I to do so, I should find it necessary to remove some of the Lemurs from their group in the highest order of the first section, and to place them in the second section."
In the succeeding year, 1844, M. Milne-Edwards, one of the most distinguished physiologists and zoologists of modern France, proposed, in a highly philosophical paper upon zoological classification in general,4 a method of subdividing the Mammalia, essentially similar to that put forward incidentally by Von Baer and Eschricht, but lacking, as I conceives what is the great merit of the latter writers, namely, the clear perception of the classificatory value of the intimate structure of the placenta and the entrance, or not, of a decidual uterine element into its composition. M. Milne-Edwards dwells with great force (as Mr. Waterhouse had previously done) upon the closeness of the general structural affinities which unite the Rodentia, Insectivora, Cheiroptera, Quadrumana, and Bimana of Cuvier together, and shows that these affinities are denoted by the discoid placenta which they possess in common.
The diffused placenta (under which head the cotyledonary placenta is included) is stated to be the characteristic of the Ruminantia, Pachydermata, Edentata, and Cetacea; while, lastly, the Carnivora and seals (Amphibies) are distinguished from all the rest by their zonular placenta."
The singular genus Hyrax, which Cuvier endeavoured to prove to be a true Pachyderm, is considered by M. Milne-Edwards to form one of the series of Mammals with a zonular placenta; and to represent, in that series, the Pachyderms in the series with diffuse placentation, and the Rodents, in the series with discoidal placentation.
M. Gervaig, in France, and M. Vogt, in Germany, have adopted the placental classification of Milne-Edwards; while, in 1807, Mr. Waterhouse's proposed, but immediately rejected, cerebral classification was substantially revived by Professor [96] Owen, in his paper "On the Characters, Principles of Division, and Primary Groups of the class Mammalia," published in the Journal of the Linnæan Society; though it should be added that Professor Owen made certain additions to the nucleus furnished by Mr. Waterhouse, which are unquestionably original.
Thus the "Lissencephala " of Professor Owen is simply a new name for the group of Mammals ("in which the cerebrum is destitute of convolutions, or nearly so") indicated by Mr. Waterhouse; and "Gyrencephalia" is a like verbal equivalent for Mr. Waterhouse's group of Mammals characterized by having the brain provided with distinct convolutions. But Mr. Waterhouse does not mention Man at all, while Professor Owen creates a new sub-class, Archencephala, for the genus Homo, and substitutes the name "Lyencephala" for Implacentalia, formerly applied to the Ornithodelphia and Didelphia.
In attempting to decide between the various classifications thus presented to us, the canons by which our judgment must be guided are simple enough. It is obvious, in the first place, that the definition of a group, whether that definition be based on cerebral or on placental characters, must be true, as a matter of fact, if any value is to be attached to the classification of which that definition forms a part.
And, in the second place, it is clear that the definition of each group must be distinctive, that is to say, it must not include the members of other groups.
Applying the second canon to the classification last mentioned, it appears to me to collapse at once.
The sub-class Lissencephala, for example, is thus defined:–
"The corpus callosum is present, but connects cerebral hemispheres as little advanced in bulk or outward character as in the preceding sub-class; the cerebrum leaving both the olfactory lobes and cerebellum exposed, and being commonly smooth, or with few and simple convolutions in a very small proportion, composed of the largest members of the group. The Mammals so characterised constitute the sub-class Lissencephala."– L. c., p. 14.
[97] On the other hand, the sub-class Gyrencephala receives the following definition:—
"The third leading modification of the Mammalian cerebrum is such an increase in its relative size, that it extends over more or less of the cerebellum, and generally more or less over the olfactory lobes. Save in very few exceptional cases of the smaller and inferior forms of the Quadrumana, the superficies are folded into more or less numerous gyri, or convolutions, whence the name Gyrencephala, which I propose for the third sub-class of Mammalia."–L.c., p. 18.
I am quite unable to see what these so-called definitions define. If, for example, we place the brains of an Ant-eater, or of a Capybara, side by side with that of a Genett–the two former being Lissencephala, the latter one of the Gyrencephala–either "definition" will apply equally well to either of the three brains. All three have slightly convoluted brains; in all three the olfactory lobes and cerebellum are more or less uncovered; and nothing in the definitions of the sub-classes of this "cerebral classification" would enable an anatomist to say that any one of these three brains belonged to one sub-class rather than another.
Since Mr. Waterhouse pointed out the fact, no one has doubted that, as a general rule, the brains of the so-called "Gyrencephala" are more convoluted, size for size, than those of the "Lissencephala;" and the relations of the size and the zoological position of an animal to the characters of its cerebral surface have long since been well discussed by Gratiolet, Dareste, and others. But it is exactly because the rule is only a general one, and has many exceptions, that the degree of cerebral convolution must be rejected as the basis of the definition of any large group of Mammals.
Thus far, we meet, in Professor Owen's definitions, with a certain foundation in fact, though it may not be such as is fitted to afford ground for classification; but the group "Archencephala" is in a more unfortunate position. Our first canon comes into operation, and we must reject it, because the statements respecting matters of fact in its definition are untrue. The words stand thus;–
[98] "In man the brain presents an ascensive step in development, higher and more strongly marked than that by which the preceding sub-class was distinguished from the one below it. Not only do the cerebral hemispheres overlap the olfactory lobes and cerebellum, but they extend in advance of the one and further back than the other. Their posterior development is so marked, that anatomists have assigned to that part the character of a third lobe; it is peculiar to the genus Homo, and equally peculiar is the 'posterior horn of the lateral ventricle,' and the 'hippocampus minor,' which characterises the hind lobe of each hemisphere."–L. c., pp. 19, 20.
These are the assertions which have been repeated over and over again during the last few years; but, thanks to the exertions of the able conservator of your Museum, it is in my power to lay before you visible and tangible facts, which prove these assertions to be wholly devoid of foundation.
The third lobe, characterized by extending farther back than the cerebellum, is said to be "peculiar to the genus Homo."
I place before you casts of the cranial cavity, accurately representing the relative positions of the parts of the brain of a Gorilla, of a Chimpanzee, of an Orang, of a Cynocephalus; and you observe that the posterior, or third lobe, of each projects further back than the cerebellum, in just the same sense as a man's can be said to do so; and in some cases, as in the baboon, to a much greater extent.
The assertion that the third lobe, as defined by Professor Owen, is "peculiar to man," is therefore demonstrably contrary to fact.
"Equally peculiar is the posterior horn of the lateral ventricle."
Side by side upon the table are two dissections, made in the same way, the one of the brain of an Orang-utan, the other that of a man, taken at hazard by Mr. Flower, who has been good enough to dissect both (Fig. 40).
Every one in this theatre, I imagine, can see perfectly well that the Orang has a posterior cornu, which, in proportion to the size of its brain, is just as long and nearly as much incurved [99] as that of the man, while it is a good deal wider at its commencement.
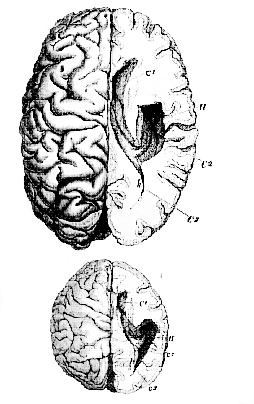
Fig. 40.–Figures [reduced to the same scale] of the dissected brains of a Man and of an Orang which were exhibited in the theatre of the Royal College of Surgeons.–
C1,
anterior cornu; C2,
descending cornu; C3,
posterior cornu; H, hippocampus major; h, hippocampus minor.
[100] In fact, even if the posterior cornu had not been demonstrated (as it has now been) in the brain of numerous genera of Apes, this one example would sufficiently demonstrate the assertion, that the posterior cornu is "peculiar to the genus Homo," to be simply untrue.
Lastly, as regards the hippocampus minor–which is also said to be "peculiar to man"–that structure is, as you perceive, as distinct in the Orang's as in the man's brain, so that the third term of the definition of the "Archencephala " is as contradictory to plain fact as the other two (Fig. 40).
Even were the posterior lobe, the posterior cornu, and the hippocampus minor peculiar to man, as supposed by the definer of the sub-class "Archencephala," instead of being, as they really are, structures far better developed in some of the lower apes than in him, their classificatory value would be extremely doubtful seeing that they are among the most variable of structures in the human brain. The casts upon the table of a Tartar's and of an Australian brain-case will demonstrate to you how insignificant may be the projection of the posterior lobe in one man and how great it may be in another. While the practical anatomists and demonstrators whom I address will be familiar with the singular variability of the posterior cornu and the hippocampus minor– structures which, without any assignable cause, or noticeable modification of the structure, or of the functions, of the brain, may present every degree of development, from absence to great size.
So little, indeed, is any zoological value to be attached to such a character as the degree of projection of the posterior lobe, that closely allied apes present us with most singular differences in this respect. Thus the group of South American monkeys which comprises the Squirrel monkey (Chrysothriæ), the posterior lobes of whose brain project beyond the cerebellum far more than they do in man, contains also the Howling monkey (Mycetes), in which the posterior lobes cannot be said to project at all. And within the last two days, Mr. Flower has discovered (and the cast upon the table enables me to demonstrate the fact to you) that in, at any rate, one species of Gibbon, the Siamang (Hylobates Syndactylus) the cerebellum projects behind the pos[101]terior lobes, while, in the three other genera of anthropoid apes, the posterior lobes of the cerebrum project behind the cerebellum.5
The latest form of the "cerebral" classification of the Mammalia having thus been shown to be devoid of any sound foundation, I proceed to inquire whether the "placental" classification does, or does not, stand upon a more secure basis, if we take, not merely, with Milne-Edwards, the form of the placenta, but with Von Baer and Eschricht, its structure, into account. It is a well-established fact that two very distinct types of placenta are to be met with in the Monodelphia, and that, at the present moment, we have no knowledge of any transitional forms between these two types. The first of these types is that exhibited by the human placenta, the second by that of the pig or horse.
From the commencement of gestation, the superficial substance of the mucous membrane of the human uterus undergoes a rapid growth and textural modification, becoming converted into the so-called "decidua." While the ovum is yet small, this decidua is separable into three portions,–the decidua vera, which lines the general cavity of the uterus; the decidua reflexa, which immediately invests the ovum; and the decidua serotina, a layer of especial thickness, developed in contiguity with those chorionic villi which persist and become converted into the fœtal placenta. The decidua reflexa may be regarded as an outgrowth of the decidua vera; the decidua serotina as a special development of a part of the decidua vera. At first, the villi of the chorion are loosely implanted into corresponding depressions of the decidua; but, eventually, the chorionic part of the placenta becomes closely united with, and bound to, the uterine decidua, so that the fœtal and maternal structures form one inseparable mass.
In the meanwhile, the deeper substance of the uterine [102] mucous membrane, in the region of the placenta, is traversed by numerous arterial and venous trunks, which carry the blood to and from the placenta; and the layer of decidua into which the chorionic villi do not penetrate acquires a cavernous, or cellular, structure from becoming burrowed, as it were, by the innumerable sinuses into which these arterial and venous trunks open. In the process of parturition, the decidua serotina splits through this cellular layer, and the superficial part of it comes away with the umbilical cord, together with the fœtal membranes and the rest of the decidua; while the deeper layer, undergoing fatty degeneration and resolution, is more or less completely brought away with the lochia, and gives place to a new mucous membrane, which is developed throughout the rest of the uterus, during pregnancy; but, possibly, arises only after delivery over the placental area.
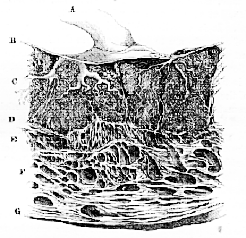
Fig. 41 .–Section of the Human Uterus and Placenta at the thirtieth week of pregnancy (After Ecker.)–
A, umbilical cord; B, chorion; C, the fœtal villi separated by processes of; D, cavernous decidua; E, F, G, wall of the uterus.
In the Pig the placenta is an infinitely simpler structure. [103] No "decidua" developed; the elevations and depressions of the unimpregnated uterus simply acquire a greater size and vascularity during pregnancy, and cohere closely with the chorionic villi, which do not become restricted to one spot, but are developed from all parts of the chorion, except its poles, and remain persistent in the broad zone thus formed throughout fœtal life. The cohesion of the fœtal and maternal placenta, however, is overcome by slight maceration or post-mortem change; and, at parturition, the fœtal villi are simply drawn out, like fingers from a glove, no vascular substance of the mother being thrown off.
The process by which the mucous membrane of the uterus returns to its unimpregnated condition after parturition in the pig has not been traced.
The extreme cases of placentation exhibited by man and by the Pig may be termed, with Von Baer and Eschricht, from the character of the maternal placenta, "caducous" and "non-caducous,'" or, from the degree of cohesion of the two placentæ in parturition, "coherent" and "incoherent;" or, what perhaps would be better still, the two Mammals may be spoken of as "deciduate" and "non-deciduate."6 But, whatever terms be employed, the question for the classifier is to inquire what mammals correspond with Man and what, with the Pig, and whether the groups of deciduate and non-deciduate Monodelphia thus formed, are natural groups, or, in other words, contain such orders as can be shown, on other grounds, to be affined.
With respect to the deciduate Monodelphia, it is certain that the apes agree, in the main, with man in placental, as in other important characters; and, so far as has hitherto been observed (though our knowledge of the placentation of the Lemurs is very defective), their placentæ differ from those of Man only in presenting a more marked location–a character which occurs as a variety in Man.
[104] The Cheiroptera, Insectivora, and Rodentia agree with Man in possessing a placenta which is not only as much "discoidal," allowance being made for the shorter curve of the uterine walls, as his, but also entirely resembles his in being developed in conjunction with a decidua. This decidua always corresponds to at least. the decidua serotina of Man; frequently there is a well-developed decidua reflexa.7 How far a decidua vera can be said to be developed is doubtful.
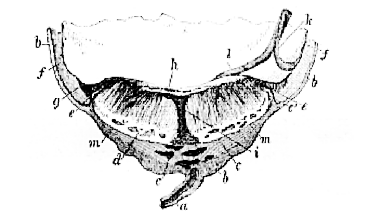
Fig. 42.–Magnified view of a section of the placenta and uterus of a pregnant Rat.
I am well aware that these statements are in direct opposition to some that have been very confidently put forward. Thus, Professor Owen, in arguing against the views entertained by Milne-Edwards and Gervais, makes the following assertions:–
"The degree of resemblance in outward form between the placenta of the Rat or Hare, on the one hand, and the Mycetes and Macacus on the other, seems to me to be more than counterbalanced by the difference of structure. The pedunculate and cotyloid placenta of the Rat consists of fœtal parts exclusively; the maternal areolar portion is as distinct from it as it is in the cotyledon of the Ruminant, and is a persistent structure of [105] the uterus. The discoid placenta of the monkey includes a large proportion of maternal cellular structure, which comes away with the fœtal portion. The difference in the organic interblending of the circulatory organs of mother and offspring, between the Rodentia and Quadrumana, is of much more real importance than the degree of superficial similarity."–L. c., p. 16, note.
Led by the extraordinary contradictions of some of the best-known facts of embryology8 contained in the passage [106] I have italicised to look into the matter afresh, I have found that the assertions made therein respecting the placenta of the Rat are as completely contrary to fact as are those respecting the brain of the apes which I have already cited from the same author.
Figure 42 represents a section of the uterus, chorion, and partially-injected placenta of a fœtal Rat, one inch and a quarter long, taken in a direction perpendicular to the long axis of the uterine cornu. a is the mesometrium traversed by a large uterine vein; b is the wall of the uterus becoming looser in texture and traversed by large venous channels in its inner substance, c; d is a decidual layer of the uterus of a cavernous structure, whence vascular processes are continued towards the chorionic surface of the placenta. A large vein (i) passes directly from the decidual layer (d), and the uterine sinuses beneath it, to near the chorionic surface of the placenta, beneath which it branches out horizontally. The chorion (f), rendered vascular over its non-placental part by the omphalomeseraic vessels (k), only begins to exhibit villous processes and folds at the point (g). These outermost villi appear to me to be free; but, more internally, they become closely connected with the upper surface of the placenta; and over the central third of the fœtal face of the placenta, the umbilical vessels (l) ramify in a radiating fashion, and send prolongations [107] down between the decidual lamellæ. The slightest traction exerted upon the cord causes the placenta to separate along the line e, m, m, e, bringing with it, of course, the cup-shaped decidua, d.9
It is obvious, from the above description, that the "pedunculate and cotyloid" placenta of the Rat does not "consist of fœtal parts exclusively," but that, on the contrary, as Eschricht has so well pointed out, "the organic interblending of the circulatory organs of mother and offspring" is as complete in the Rat as in Man; and that, therefore, the concluding paragraph of the citation from Professor Owen's paper ought to be reversed.
The Carnivora develop a well-marked decidua, and their [108] placenta in all genera which have been examined (except the Polecat, according to Von Baer) has the form of a complete zone, or broad girdle, surrounding the middle of the chorion and leaving the poles bare (Fig. 43).
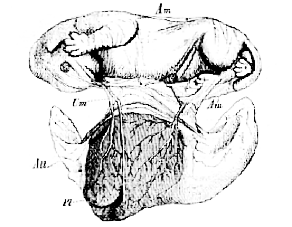
Fig. 43.–Fœtal kitten, with its membranes and placenta. The latter is seen from within, the chorion and allantois being open and everted.–
Am, amnion; All, allantois; Pl, placenta; Um, umbilical vesicle. (From a preparation in the Museum of the Royal College of Surgeons.)
Thus Man; the Apes, or so called Quadrumana; the Insectivora; the Cheiroptera; the Rodentia, to which the lowest apes present so many remarkable approximations; and the Carnivora (united into one group with the Insectivora by Cuvier) are all as closely connected by their placental structure as they are by their general affinities.
With the Pig, on the other hand, all the Artiodactyla, all the Perissodactyla (save one, taking the group in its ordinarily received sense) and all the Cetacea which have been studied, agree in developing no decidua, or, in other words, in the fact that no vascular maternal parts are thrown off during parturition. But considerable differences are observed in the details [109] of the disposition of the fœtal villi, and of the parts of the uterus which receive them. Thus, in the Horse, Camel, and Cetacea the villi are scattered, as in the Pig, and the placenta is said to be diffuse; while, in almost all the true Ruminants, the fœtal
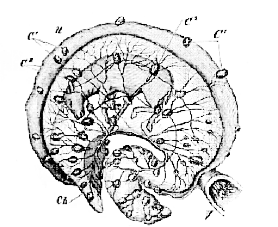
Fig. 44.–Uterus of a Cow in the middle of pregnancy laid open.–
V. vagina; U, uterus; Ch, chorion; C 1, uterine cotyledons; C 2, fœtal cotyledons (after Colin).
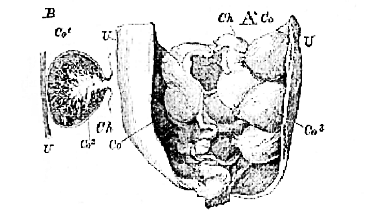
Fig. 45.–A. Horn of the Uterus of a pregnant Ewe, laid open to show,
Ch, the chorion; with Co, the cotyledons. B. Diagrammatic section of a Cotyledon.–U, uterine wall; Co1,
uterine cup of the cotyledon; Co2, chorionic villous tuft of the cotyledon. (From a preparation in the Museum of the Royal College of Surgeons.)
[110] villi are gathered into bunches, or cotyledons, which in the Sheep (Fig. 46) are convex, and are received into cups of the mucous membrane of the uterus; while in the Cow, on the contrary, they are concave, and fit upon corresponding convexities of the uterus (Figs. 44 and 46). . . . .
No one, probably, would be inclined to object to the association of the orders just mentioned into one great division of the Monodelphia, characterised by its placental structure. But such grouping leaves several important points for discussion. The Elephant, as Professor Owen10
has shown, has a zonary placenta, and the genus Hyrax has been known since the time of Home to be in like case. Hence, as the elephants are commonly supposed to be closely allied with the Pachydermata, which possess diffuse, non-deciduate placentas and as Hyrax is now generally, if not universally, admitted into the same order as the Horse, which has a diffuse, non-deciduate placenta, it is argued that placental characters do not indicate natural affinities. A question, indeed, arises, which has not been answered by those who have described the placenta of Elephas and Hyrax. Is the placenta of these animals simply a zone-like arrangement of villi [111] or cotyledons, in connection with which no decidua is developed, or is it a true deciduate placenta, resembling that of the Carnivora in the essentials of its internal structure as in its external form? Recent investigation has convinced me, that, in both these animals, the placenta is as truly deciduate as that of a Rodent; so that most unquestionably, if the placental method of classification is to be adopted, both Elephas and Hyrax must go into the same primary division of the Monodelphia as the Rodentia and Carnivora.
But do these facts really present obstacles to the placental system of classification?
So far as the case of the Elephants is concerned, I must confess that I see no difficulty in the way of an arrangement which unites the Proboscidea more closely with the Rodentia than with the Artiodactyla and Perissodactyla, the singular ties which unite the Elephants with the Rodents having been a matter of common remark since the days of Cuvier.
In the absence of any definite knowledge of the placental structure of Rhinoceros and Tapirus,11
it would, perhaps, be premature to discuss the position of Hyrax, as determined by its placenta; but if it should eventually appear, as is very probable, that Rhinoceros, like Tapirus and Equus, has a diffuse, nondeciduous placenta, I should have no hesitation in regarding Hyrax as the type of a distinct order of deciduate Monodelphous Mammalia. Hyrax, in fact, hangs by Rhinoceros mainly by the pattern of its molar teeth,–a character which affords anything but a safe guide to affinity in many cases.12
Concerning the placentation of the Sirenia we have no information.
Among the Edentata, the Sloths have presented a cotyledonary placenta, and the Armadillos have been affirmed to possess a discoidal one. I am not aware that the minute structure of the placenta has been examined in either of these groups, but I am [112] indebted to Dr Sharpey for valuable information respecting the placental structure of Manis. The surface of the chorion is covered with fine reticulating ridges, interrupted here and there by round bald spots, giving it an alveolar aspect, something like the inside of the human gall-bladder, but finer. The inner surface of the uterus exhibits fine low ridges or villi, not reticulating quite so much. The chorion presents a band, free from villi, running longitudinally along its concavity, and there is a corresponding bald space on the surface of the uterus. The ridges of the chorion start from the margins of the bald stripe, and run round the ovum. The umbilical vesicle is fusiform. This is clearly a non-deciduate placenta, and the cotyledonary form of that of the Sloth leads me to entertain little doubt that it belongs to the same category.
Admitting all these difficulties and gaps in our information, it still appears to me that the features of the placenta afford by far the best characters which have yet been proposed for classifying the Monodelphous Mammalia, especially if the concomitant modifications of the other fœtal appendages, such as the allantois and yelk-sac, be taken into account. And it must be recollected that any difficulties offered by the placental method attach with equal force to the systems of classification based upon cerebral characters which have hitherto been propounded. If any objections, on the ground of general affinities, are offered to the association of Elephas, Hyrax, Felis, and Cercopithecus in the same primary mammalian division of deciduate Monodelphia, they are not removed by constructing that primary division upon other principles, and calling it Gyrencephala.
1 For a number of years I have entertained the gravest doubts respecting the accuracy of the doctrine put forth now nearly thirty years ago by Professor Owen, and almost universally received, that the corpus callosum is absent in Monotremes and Marsupials, and at one time I began to collect materials for the thorough investigation of the question; but other occupations intervened, and the plan was never carried out. Nevertheless, I have always expressed myself cautiously on this subject, and, as the text shows, I was particularly guarded when delivering the present lecture. At that time, in fact, I was well aware that my friend Mr. Flower had commenced a series of inquiries into the question, and such results as he had then obtained tended greatly to the increase of my skepticism. Mr. Flower has since been good enough to go carefully with me over the large series of drawings and preparations which he has made; and I am prepared to express my entire concurrence in his conclusion that the corpus callosum exists, distinctly developed, though not so well as in monodelphous, or placental, Mammals, in both the Didelphia and the Ornithodelphia.
2 "Comparative Anatomy," vol. iii
3 "Annals and Magazine of Natural History," 1843, vol. xii. p. 399.
4 "Annales des Sciences naturelles." Serie 3. Tome 1. "Considerations sur quelques Principes relatifs à la Classification naturelle des Animaux."
5 See Mr. Flower's paper "On the Brain of the Siamang," Natural History Review, April, 1863. "This peculiarity of the Siamang's brain is due to two causes–firstly, the large development of the cerebellum; secondly, and I shall afterwards show, mainly, to the actual shortness of the posterior or occipitai lobe of the cerebrum."–L.c., p. 282.
6 It is, of course, by no means intended to suggest by these terms, that the homologue of the decidua does not exist in the "non-deciduate" Mammals. The mucous membrane of the uterus becomes hypertrophied during pregnancy in both the deciduate and the non-deciduate Mammals; but it is thrown off, and so gives rise to a "decidua" only in the one of these two groups.
7 See upon this subject the recently-published valuable essay of Reichert: "Beitrage zur Entwickelungs-geschichte des Meerschweinchens." Reichert finds a complete, or almost complete, decidua reflexa in Rats, Mice, Guinea-pigs, and Bats; while in Rabbits, Hares, and Carnivora, the decidua reflexa only partially surrounds the ovum.
8 Eschricht described the placenta of the Bat with great precision, as the following extract will show, six-and-twenty years ago. Is it possible that a hasty perusal of a passage which I have put into italics below should have misled any of his successors into supposing that Rodents have persistent cotyledons like those of Ruminants?–[.... 106] "Gliribus si ut ruminantibus pars uterina placentæ in partu non abstruderetur, mirum sine fuissel...."
9 My friend Professor Rolleston has made the following statements in a paper which will shortly appear in the Zoological Society's Transadions.
1. The Rat's afterbirth consists of a saucer-shaped deciduous serotina, and a button shaped placenta proper. Afterbirths made up of these two elements may be found in the stomachs of animals of this species after parturition, as they, like many other Mammals below the Simiadæ, devour them. Under these circumstances, the two constituent factors of the afterbirth may either be found in their normal connection or they may be separated one from the other.
2. The non-deciduous part of the serotina forms in the Rat, after parturition, a hernial protrusion into the mesometrium, which has been mistaken for a developing ovum (see Hunterian Catalogue, Phys. Ser. Prep. 3466); just as the homologous structures in the human subject form a hernial protrusion into the cavity of the uterus, which may persist as a more or less elevated area for several years. (Cf. Robin, Mem. Imp. Acad. Med., tom. xxv., p. 137.)
3. The homologue of the saucer-shaped deciduous serotina of the Rat is, in the human subject, the thin layer of laminated albuminous tissue, which, in a placenta expelled without suffering much violence, is seen clothing its uterine surface. It is smaller, relatively, to the other structures concerned in the nutrition of the fœtus, in the human than in any other species. It is more easily demonstrable in the Monkey (Macacus, Nemestrinus, e. g.), as being a more coherent and stouter membrane than in Man. It is, however, here still a condensed and membranous structure, as compared with its pulpy homologues in Carnivora, Insectivora, and Rodents.
4. In early periods of utero-gestation in the common Shrew and Hedgehog, the deciduous serotina is a very much larger structure than the placenta proper, which it entirely covers, except on the fœtal aspect. But in the Tenrec, near the full time, the decidua serotina is of but wafer-thickness.
5. Dr. Matthews Duncan and M. Robin have shown that the muscular coat of the uterus is never left denuded after parturition in the human subject. The same remark holds good in the case of the "deciduate" Mammalia, in all of which a more or less modified mucous tissue, the "non-deciduous serotina," is left, after parturition, upon the utero-placental area, from which the deciduous serotina and placenta proper have been separated as "afterbirth."
10 Description of the Fœtal Membranes and Placenta of the Elephant."
Philosophical Transactions, 1857.
11 Home's description of the fœtal membranes of the Tapir is very poor, but Bauer's beautiful figures show clearly that the villi are diffuse, as in the Horse.
12 See, in reference to this point, the late Professor A. Wagner's excellent remarks on Cuvier's exaggeration of the Rhinocerotic affinities of Hyrax, in Schreber's "Säugethiere." Supp. Band. Abth. iv. p. 307.
[113]
Lecture 7. On the Vertebrate Skull.
The Structure of the Human Skull.
The human skull is by no means one of the simplest examples of a vertebrate cranium which can be studied, nor is the comprehension of its structure easy; but, as all vertebrate anatomy has started from the investigation of human organization, and the terms osteologists use are derived from those which were originally applied to definite parts of the organism of man, a careful investigation of the fundamental structure of man's skull, becomes an indispensable preliminary to the establishment of anything like a sound comparative nomenclature, or general theory, of the Vertebrate Skull.
Viewed from without (Fig. 47), the human cranium exhibits a multiplicity of bones, united together, partly by sutures, partly by anchylosis, partly by moveable joints, and partly by ligaments; and the study of the boundaries and collections of these bones, apart from any reference to the plan discoverable in the whole construction, is the subject of the topographical anatomist, to whom one constantly observed fact of structure is as valuable as another. The morphologist, on the other hand, without casting the slightest slur upon the valuable labours of the topographer, endeavours to seek out those connections and arrangements of the bony elements of the complex whole which are fundamental, and underlie all the rest; and which are to the craniologist that which physical geography is to the student of geographical science.
[114] Perhaps no method of investigating the structure of the skull conduces so much towards the attainment of a clear understanding of this sort of architectural anatomy, as the study of sections, made along planes which have a definite relation to the principal axes of the skull.
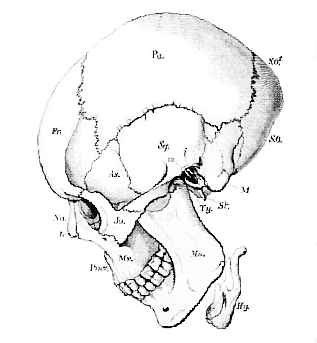
Fig. 47.–Diagrammatic side view of a Human Skull.–
Fr. Frontal. Pa. Parietal. S. O. Supra-occipital. S.O1. Squama occipitis above the torcular Herophili and lateral sinuses. As. Alisphenoid. Sq. Portio spumosa of the temporal bone. M. Mastoid process and pars mastoidea. Ty. Tympanic. St. Styloid process. Na. Nasal. L. Lachrymal. Ju. Jugal, or Malar. Pmx. Premaxilla. Mx. Maxilla. Mn. Mandible. Hy. Hyoid. m. Malleus. i. incus. [Three letters will bear the same signification throughout the series of figures of crania.]
[115] If a vertical and transverse section be taken through the cranium, in such a manner that the plane of the section shall traverse both external auditory meatuses, the skull will be divided into two unequal portions–an anterior, larger, and a posterior, smaller. The former, if viewed from behind, will present the appearance represented in Fig. 48.
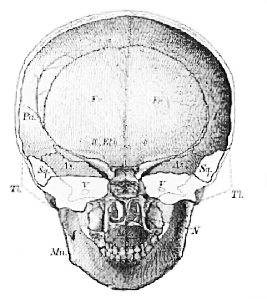
Fig. 48.–Anterior half of the skull of a young person (six or seven years of age) transversely bisected.
The temporal bone (Ti) on each side is left in outline, and the contour of the alisphenoid is supposed to be seen through it.–II, optic foramina between the roots of the orbito-sphenoid; V, foramen ovale for the third division of the trigeminal; N indicates the nasal chamber; Mx is placed in the buccal chamber.
A stout median floor (BS) whence lateral continuations (AS) are prolonged to meet an arched roof (Pa), divides a capacious upper chamber, which, during life, lodged a part of the brain, from a lower chamber, formed by the bones of the face. This lower chamber itself is again separable into two [116] parts,–an upper, divided into two by a median septum, the nasal passages; and a lower, the oral cavity.
The posterior portion of the bisected skull (Fig. 49) presents, in like manner, a strong floor (BO) and a large upper chamber for the lodgment of parts of the brain; but the lower chamber seems at first to be absent in the skeleton, being represented, in fact, only by the styloid processes (St), the so-called stylo-hyoid ligaments, and the hyoidean bone (Hy) which is suspended by these ligaments to the skull.
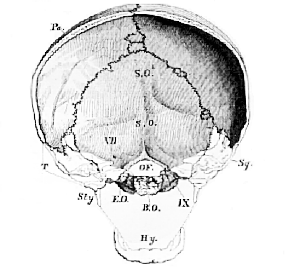
Fig. 49.–The posterior half of the transversely bisected skull, Fig. 48.–
B.O., the basi-occipital, E.O., E.O., the ex-occipitals, T, the temporal bone left in outline; O.F., occipital foramen; VII. canal for the portio dura and portio mollis; I.X., foramen for the ninth or hypoglossal nerve.
A longitudinal and vertical section of the skull (Fig. 50) enables us to observe the same relations of the parts from another point of view. The central bones (BO, BS, PS, Eth., Vo), which lie between the arches of the brain-case above, and the arches of the face below, are, in such a section, found to [117] constitute a continuous series, from the occipital foramen to the anterior extremity of the nasal passages, which, as it forms the common centre or axis, not only for the bones of the brain-case or cranium proper, but also for those of the face, may be termed the Cranio-facial axis.
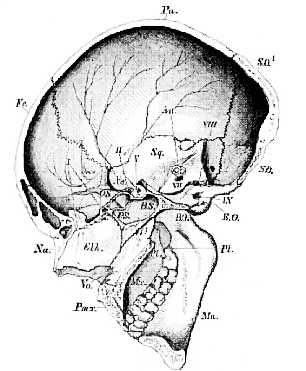
Fig. 50.–Longitudinal and vertical section of a Human skull.–
* The sella turcica. Au. The position of the superior and posterior vertical semicircular canals. I., II., V., VIII., IX. The exit of the olfactory, optic, third division of the fifth, eighth, and ninth nerves. Vo., the Vomer.
It will be useful to divide this axis into two portions,–a [118] posterior basi-cranial (BO, BS, PS), which forms the centre of the floor of the proper cranial cavity; and an anterior, basi-facial (Eth., Vo.),which constitutes the axis of the front part of the face.
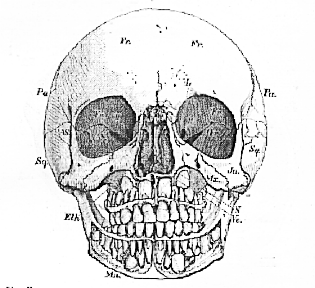
Fig. 51.–Front view of the skull, the halves of which are shown in Figs. 48 and 49.–
N. nasal chamber; Or, orbit. The nasal bones are removed, and so much of the upper and lower jaws as is necessary to show the permanent teeth.
Three pairs of chambers, destined for the lodgment of the organs of the higher senses, are placed symmetrically upon each side of the double bony box thus described. Of these, two pair are best seen in a front view of the skull (Fig. 51), the inner pair being the olfactory, or nasal chambers (N), the outer pair, the orbits (Or). The other pair are better displayed in the transverse sections, Fig. 48 and Fig. 49, and are formed by the temporal bones of anatomists (T. Tl), and especially by the petrous and mastoid portions of those bones.
There is an obvious difference between the relations of these sensory chambers to the contained sensory organ, in two of [119] these chambers as compared with the third. The sensory apparatuses of the nose and of the ear are firmly fixed to, or within, the bony chambers in which they are lodged. That of the eye, on the other hand, is freely moveable within the orbit.
An axis, upper and lower arches, chambers for the sensory organs–such are, speaking generally, the components of the skull The special study of these components may be best commenced from the cranio-facial axis. Viewed either from above (Fig. 52) or from below (Fig. 53j, the cranio-facial axis is seen to be depressed, or flattened from above downwards, behind, and thick and nearly quadrate in the middle; while, in front, it is so much compressed, or flattened from side to side, that it takes the shape of a thin vertical plate. In such a young skull as that from which the Figures 52 and 53 are taken, the depressed hindermost division of the axis is united with the rest, and with the bones EO, EO, only by synchondroses; and is readily separable, in the dry skull, as a distinct bone, which is termed the Basi-occipital (BO). This basi-occipital furnishes the front boundary of the occipital foramen; and its postero-lateral parts, where they abut against the bones EO, contribute,
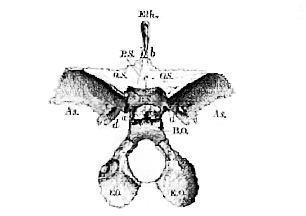
Fig. 52–Cranio-facial axis and lateral elements of the superior arches of a human skull viewed from above.–
a, the spheno-occipital synchondrosis; b, the ethmo-sphenoid synchondrosis; c, the tuberculum sellæ, indicating the line of demarcation between the basi-occipital and the presphenoid; d, the lingulæ sphenoidales.
[120] to a small extent, to the formation of the two occipital condoles. In the adult skull the basi-occipital anchyloses completely with the ex-occipital. on the one hand, and with the next bone of the basi-cranial axis on the other, so that the saw must be called to our aid in order to demonstrate the bone. . . .
From the synchondrosis a to the point b, in even so young a skull as that here represented, the basi-cranial axis is formed by one continuous ossification, the basi sphenoid bone, excavated superiorly (Figs. 50 and 52) by a saddle-shaped cavity, the sella turcica, which lodges the pituitary body,–an organ of no great physiological moment, so far as we know, but of first-rate morphological significance.
On each side of the hinder part of the sepia turcica, the basi-sphenoid presents a groove for the internal carotid artery, and this groove is completed in front and externally, by an osseous mass, tapering from behind forwards, the lingula sphenoidalis, which lies between the basi-sphenoid and alisphenoid. At the front part of the sepia, separating it from the [121] depression for the optic commissure, there is a transverse ridge, the tuberculum sellæ.
. . . .
[126] In like manner, the face may be resolved into a series of bones, occurring in pairs from before backwards, and forming more or less well-defined lower arches, some of which embrace the nasal cavity, being placed in front of, or above, the oral aperture, while others enclose the buccal chamber, and are situated behind and below the oral aperture. Of the former, pre-oral bones, there are four pairs–-the Premaxillæ (Pmx.), the Maxillæ (Mx.), the Palatines (Pl.), and the Pterygoids (Pt.).
The Premaxillæ, which lodge the upper incisor teeth, so early lose their distinctness in man, by becoming anchylosed with the maxillary hones (at any rate externally and anteriorly) that they are rarely recognised as separate bones. Nevertheless, suture extending upon the bony palate from the posterior margin of the alveolus of the outer incisors to the incisive foramen, very commonly persists, as an indication of the primitive separation of these bones. The most important character of the premaxillæ, regarded morphologically, is, that they are connected, superiorly, with the anterior termination of the cranio-facial axis, and that this connection is a primary one. Each premaxilla passes from its inner end, which is united with the axis, outwards and backwards, and two of the other three pair of pre-oral bones have similar relations to the cranio-facial axis. The anterior of these are the Palatine bones; the inner, or sphenoidal, processes of which are connected with the basi-[126]sphenoid and with the vomer; while the outer, or orbital, processes articulate with the so-called lateral masses of the ethmoid and with the maxilla; so that the upper part of each palatine bone is directed, from the cranio-facial axis, with which its inner end is connected, outwards and forwards (Fig. 54). The third pair of bones, the Pterygoids, are the internal pterygoid processes,–bones which are originally quite distinct from the sphenoid, while the external pterygoid processes are of a very different character, being mere outgrowths of the alisphenoids. These are connected with the basi-sphenoid (or rather with the lingulæ sphenoidales), above, and, in front, with the palatines, while their planes are directed backwards and somewhat outwards. The fourth pair of pre-oral bones–the Maxillæ–are connected in front and internally with the premaxillæ, and behind and internally with the palatines, but they nowhere come into direct contact with the cranio-facial axis, at least primarily.
I make the latter qualification because the vomer articulates with the superior surface of the palatine plates of the maxillæ, and it may be said that, in this way, the maxillæ do unite with the cranio-facial axis. This articulation, however, has nothing to do with the primitive connections of the bones, but depends upon a modification of the maxillæ peculiar to the higher vertebrata. The bony apertures–called "posterior nares"–in Man, for example, are structures of a totally different character from, and superadded to, what are called the posterior nares in a frog, or ordinary lizard, or bird. In these lower Vertebrates, the posterior nares are apertures, bounded on the inner side, by the vomer; on the outer side and behind, by the palatine bones; in front, by the premaxillæ and maxillæ. In Man, on the other hand, the apertures so called are limited, it is true, on the inner side by the vomer, and on the outer side by the palatine bones; but they are also bounded below and in front by the palatine bones, and the premaxillæ and maxillæ have nothing to do with them. On looking closely into the matter, however, it will be found that the region of the palatine which forms the outer and inferior boundary of the posterior nares of Man is a something which has no representative in the lower Vertebrate.
[127] But if, with a fine saw, the greater part of the perpendicular plate of the palatines, and the corresponding part of the maxillaries, and, with these, their palatine plates, be cut away, leaving only the premaxillæ, vomer, and upper parts of the maxillary and palatine bones; it will be found that hinder nares are left, which entirely correspond with the "posterior nares" of a bird or of an amphibian; that is to say, they are passages between the vomer in the middle line, the premaxillæ and maxillæ in front and externally, and the palatines externally and behind.
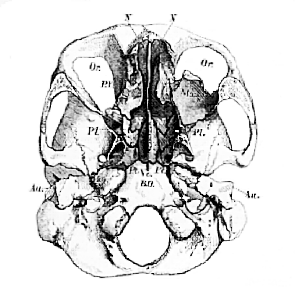
Fig. 54.–The base of a human skull–
the nasal, ethmoid, vomerine, maxillary, palatine, and pterygoid bones being cut through horizontally, and their lower portions removed. The entire right maxilla is taken away. The posterior pair of letters, NN, are situated in the median nares, which are incomplete, in front, in consequence of the removal of the premaxillæ.
In fact, the apertures of the nasal chamber into the mouth, thus artificially exposed, are those which originally exist in Man and the higher Vertebrata; but the downward growth of the maxilla into its alveolar process, and of the palatine bone into its perpendicular plate, together with the production inwards [128] of the palatine plates of these bones, which eventually unite with the vomer, give rise to the apertures, which are ordinarily called posterior nares. So that in Man, for example, there are three pairs of "nares:"–the external, situated between the anterior end of the internasal septum, the nasal bones, and the premaxillæ, as in the lower Vertebrates; the median, between the vomer, the palatines, and the premaxillæ, which correspond with the posterior nares of the lower Vertebrates; and the posterior, between the vomer, internally, and the palatines above, at the sides, and below, which are peculiar to the higher Vertebrates.
And, to return to the maxilla, we find that it really differs altogether from the other pre-oral bones, and is, as it were, fastened on to the outer sides of the premaxillary and palatine bones, without having any primary direct connection with the cranio-facial axis.
The post-oral bones surround the buccal cavity, and form two distinct arches–the mandibular and the hyoidean. Neither of these arches is directly connected with the cranio-facial axis, nor with the segments of the brain-case, but both are suspended to different parts of the temporal bone, which is so singularly intercalated between the middle and posterior of those segments.
The lower jaw or Mandible (Mn) consists of two rami, anchylosed at the symphysis, and each consisting of a single piece, the condyle of which articulates with the squamosal.
The Hyoid hone (Hy), composed of its body and two pairs of cornua, does not articulate directly with the temporal bone, but ligaments connect it with the styloid processes, and these last bones unite with the posterior part of the periotic capsules.
Thus, the natural connections of the bones by no means allow of the separation of the walls of the lower chambers of the human skull into a series of arches springing from, and corresponding with, the axial parts, as we found to be the case with the walls of the upper chambers.
If the temporal bone be detached, the hyoidean and mandibular arches come with it, and exhibit no connection with the occipital or the parietal segments. Indeed, the latter is [129] preoccupied by the pterygoid and the palatine, both of which are connected with the basi-sphenoid (at least with the lingulæ), while the anterior part of the palatine is also connected, in the adult state, with the presphenoid, by the intermediation of the corona sphenoidalia.
Two bones yet remain to be mentioned which come neither into the category of axial bones, nor of superior or inferior arch bones, nor, strictly speaking, of sense-capsule bones. These are the Lachrymal (L), intercalated between the nasal, maxillary, and lateral mass of the ethmoid, and serving to lodge the conduit which places the orbit and the nasal cavity in communication; and the Jugal or Malar (Ju), which connects the bones of the orbital chamber with the squamosal element of the temporal bone.
The skull, thus composed, serves as a protection to the organs which are lodged within it, and which are of as great importance in their morphological, as in their physiological, aspect.
The cerebral hemispheres and cerebellum, with their dependent parts, fill the cranial cavity, the lower lateral margin of the posterior cerebral lobes corresponding with the torcular Herophili and the lateral sinuses, on the inner surface of the occipital bone; or, in other words, with the line of attachment of the tentorium. Certain axial parts of the brain have definite relations to the axial parts of the cranium. Thus, the medulla oblongata lies upon the basi-occipital. The pituitary body rests upon the upper surface of the basi-sphenoid, this bone constituting the chief part of the front as well as of the hinder wall of the sella turcica. The chiasma of the optic nerves rests upon the hinder portion of the upper face of the presphenoid, and the peduncles of the olfactory nerves upon the front portion of that face. The termination of the axial parts of the brain in the lamina terminalis of the third ventricle corresponds pretty nearly with the termination of the basi-cranial axis in the anterior extremity of the presphenoid.
Not less important are the relations of many of the cerebral nerves to the lateral elements of the arches of the brain-case.
The filaments of the olfactory nerves pass out through the [130] cribriform plates, leaving the ethmoid proper, or lamina perpendicularis, upon their inner side, and the lateral masses of the ethmoid, or superior and middle spongy bones, upon their outer sides.
The optic nerves pass out through the optic foramina, situated between the roots of the orbito-sphenoids, from the chiasma, which rests, as has just been stated, upon the posterior and upper part of the presphenoid. Hence it follows, that the presphenoid lies in front of, and between, the optic nerves, which embrace it, as in a fork, from behind.
The third and fourth pairs are not of so much morphological importance that I need dwell upon them, but the trigeminal affords first-rate cranial landmarks by its nasal branch and its whole third division. The nasal nerve enters the orbit by the foramen lacerum anterius, passes to the inner side of the eye, and then, traversing the anterior of the two ethmoidal-foramina, perforates the "lateral mass of the ethmoid," and entering the cavity of the bony cranium, though it always lies beneath the dura mater, skirts the olfactory aperture, and passes out into the nasal cavity, by an aperture in the front part of the cribriform plate. We shall find this irregular perforation of the "lateral mass of the ethmoid," by the nasal division of the fifth nerve, to be an excellent guide to the determination of the homologies of the bone in the lower Vertebrata.
The third division of the trigeminal traverses the foramen ovale in the posterior part of the alisphenoid, so that it makes its exit behind the greater part of that bone, and altogether in front of the periotic bone.
The portio dura enters the internal auditory foramen in the periotic mass, runs along its canal, situated above the fenestra ovalis, and eventually passes out by the stylo-mastoid foramen. It therefore perforates the fore part of the periotic, passing in front of the membranous labyrinth. The portio mollis also enters the periotic bone by the internal auditory foramen, and it terminates in the membranous labyrinth.
The eighth pair passes out through the foramen lacerum posterius completely behind the periotic (which thus lies between the exits of the fifth and of the eighth pairs), and in front of the exoccipitals.
[131] The ninth pair perforates the ex-occipitals in front of the condyles.
With regard to the relations of the nerves to the inferior arches of the skull, only one circumstance calls for particular notice,–the distribution of the terminal divisions of the porno dura. This nerve divides, as it is about to leave the temporal bone, into two portions, the larger of which passes out by the stylo-mastoid foramen, and, besides giving off many other branches, supplies certain muscles of the hyoidean apparatus.
The smaller division of the nerve, of comparatively insignificant size–the chorda tympani–returns to the tympanic cavity, crosses it, and leaving it by an aperture internal to, and above the tympanic element, runs down upon the inner side of the lower jaw. In Man, the great development of the facial muscles gives a predominance to the branches of the portio dura which supply them; but, in the lower Vertebrates, the nerve becomes more and more completely represented by simple mandibular and hyoidean divisions, corresponding respectively with the chorda tympani and the branches distributed to the stylo-hyoid and digastrio.
. . . .
[136]
Lecture 8 On the Structure of the Skull
The Development of the Human Skull
AS might be expected from the nature of the case, it has not yet been possible to obtain a series of human embryos, in every stage of development, sufficiently large to enable embryologists to work out all the details of the formation of the human skull. But all higher vertebrate embryos so nearly follow one and the same type of early developmental modification, that we may reason, with perfect confidence, from the analogy of the lower-Vertebrates to man, and fill up the blanks of our observations of hunian embryos by investigations of the chick, the dog, the rabbit, or the pig.
In the chick, (See Lecture IV, pp. 64-66) the first indication of the body of the embryo is an elongated, elevated area of the blastoderm, the axis of which is traversed by a linear groove. The one end of the elongated area is wider and more distinctly raised up from the rest of the blastoderm, than the other: it is the cephalic end (Fig. 31, A, a), and the linear groove stops short of the rounded extremity of this part of the elevated area. A peculiar cellular cylinder, tapering off at each end, the notochord, is soon discerned occupying the bottom of this groove, beneath the outer, serous, or neuro-epidermic layer of the germ.
A familiar outgrowth of the convex summits of the ridges which bound the primitive groove now takes place, in that part of the embryo, which will eventually become the middle region [137] of the head; and the dorsal laminæ produced, extendnig forwards and backwards, like parapets, upon each side of the primitive groove, lay the foundations of the lateral walls, not only of the skull, but of the spinal column.
Very early, however, the boundary line between skull and spinal column is laid down, by the appearance in the substance of the bases of the dorsal lamniæ and the adjacent middle layer of the blastoderm, of the first pair of those quadrate masses of condensed tissue, the proto-vertebræ ("Urwirbel" of the German writers), which are the foundations, not only of the bodies of the vertebræ, but of the spinal muscles and ganglia. The proto-vertebræ increase in number from before backwards; and, at length, extend through the whole range of the spinal column, while none ever make their appearanee in the region which will be converted into the skull.
The edges of the dorsal lamniæ now unite, the coalescence taking place first in the middle cephalic region, and extending thence backwards and forwards; at the same time, the canal becomes separated into three distinct dilatations, or cerebral vesicles. of which the anterior is by far the most marked (Fig. 57, A I, II, III).
The rudimentary cranial cavity next becomes bent upon itself in such a manner, that the longitudinal axis of the first cerebral vesicle takes a direction at right angles to the axis of the third, and of the spinal canal generally. In consequence of' this change, the middle cerebral vesicle occupies the summit of the angulation, and becomes the most anterior point of the wliole body (Fig. 57, C, D).
The bend thus produced is the cranial flexure. It results in the division of the floor of the cranial cavity into two parts, an anterior and a posterior, which are at right angles to one another (Fig. 57, C, D, E). Hitherto, no trace of the notochord has been observed in the anterior division, that structure ending in a point behind the flexure (Fig. 57, D, E, h).
As development proceeds, the anterior cerebral vesicle becomes divided into two portions, – an anterior, the vesicle of the cerebral hemispheres (I a); and a posterior, the vesicle of the third ventricle (I). In the upper wall of the vesicle of the third [139] ventricle the rudimentary pineal gland (e) makes its appearance in the middle line. From the middle of the lower wall grows out a process, the infundibulum, terininating in a glandular appendage, the pituitary body, which last is lodged in the deep fossa situated in the floor of the anterior division of the skull, immediately in front of, and beneath, the termiation of the notochord (Fig. 57, B, D, d).
The three pairs of sensory organs appertaining to the higher senses– the nasal sacs, the eyes, and the ears,–arise as simple cæcal involutions of the external integument of the head of the embryo. That such is the case, so far as the olfactory sacs are concerned, is obvious; and it is not difficult to observe that the lens and the anterior chamber of the eye are prodnced in a perfectly similar manner. It is not so easy to see that the labyrinth of the ear arises in this way, as the sac, resulting from the involution of the integument, is small, and remains open but a very short time (Fig. 57, C, b . . . .
[142] Thus far the terms of my description are almost as applicable to the embryonic cranium of Man as to that of the chick.
The human cranium has been observed forming part of an open groove; it undergoes a flexure, and develops visceral arches altogether similar to those of the chick, nor is there any reason to doubt that the organs of sense are developed in the same manner. The very earliest condition of the cartilaginous cranium of the human embryo has not been observed; but, at the beginning of the second month, it consists wholly of cartilage and of membrane, disposed in a manner which differs only in detail from that seen in the chick. Thus the occipital foramen is surrounded by cartilage, continuous with that which extends through the basi-sphenoidal, presphenoidal, and ethmoidal regions to the anterior end of the face. The alisphenoids and orbito-sphenoids are represented by cartilage, and cartilaginous plates arch down from the summit of the internasal cartilage, on each side, to form the substratum on which, the nasal bones and, in which, the spongy bones will be developed. That part of the cranial cartilage which lodges the auditory organ is exceedingly large, and constitutes, not only an oval capsule for the membranous labyrinth, but sends back a continuation which fills the space corresponding to the pars mastoidea, and extends somewhat higher than it beneath the parietal region of the skull. All the upper part of the cranium is and remains simply membranous.
[143] The relations of the regions of the chondro-cranium thus formed to the parts of the brain and to the exits of the nerves are the same as those which are observed in the bones which they prefigure.
When these bones begin to be developed, some of them make their appearance in the cartilage of the embryonic skull, some in the perichondrium, others in the membranous roof which is continuous with the perichondrium.
Fig. 58.–Successive embryonic conditions of the human head (after Ecker).–
I, II, the first and second cerebral vesicles. 1, 2, 3, 4, the visceral arches; a, the maxillary process; b, the eye; nf, the middle naso-frontal process; c, the lateral naso-frontal process; i, the tongue; au, the outer part of the first visceral cleft, which effectually gives rise to the external auditory meatus. A, at three weeks. B. at five weeks. C, at six weeks. D, at seven weeks. E, at eight weeks. F, outline side view of E.
. . . .


{kind=link}