
A Manual of the Anatomy of Invertebrated Animals (1877)
by T. H. Huxley
by T. H. Huxley
[3]
THE present volume on the Anatomy of Invertebrated animals fulfills an undertaking, to produce a treatise on comparative anatomy for students, into which I entered two-and-twenty years ago. A considerable installment of the work, relating wholly to the Invertebrata, appeared in the Medical Times and Gazette for the years 1856 and 1857, under the title of "Lectures on General Natural History." But a variety of circumstances having conspired, about that time, to compel me to direct my attention more particularly to the Vertebrata, I was led to interrupt the publication of the "Lectures" and to complete the Vertebrate half of the proposed work first. This appeared in 1871, as a "Manual of the Anatomy of Vertebrated Animals."
A period of incapacity for any serious toil prevented me from attempting, before 1874, to grapple with the immense mass of new and important information respecting the structure, and especially the development, of Invertebrated animals, which tile activity of a host of investigators has accumulated of late years.
That my progress has been slow will not surprise any one who is acquainted with the growth of the literature of animal morphology, or with the expenditure of time involved in the attempt to verify for one's self even the cardinal facts of that science; but I have endeavored, in [4] the last chapter, to supply the most important recent additions to our knowledge, respecting the groups treated of in those which have long been printed.
When I commenced this work, it was my intention to continue the plan adopted in the "Manual of the Anatomy of Vertebrated Animals," of giving a summary account of what appeared to me to be ascertained morphological facts, without referring to my sources of information. I soon found, however, that it would be inconvenient to carry out this scheme consistently; and some of my pages are, I am afraid, somewhat burdened with notes and references.
I am the more careful to mention this circumstance as, had it been my purpose to give any adequate Bibliography, the conspicuous absence of the titles of many important books and memoirs might appear unaccountable and indeed blameworthy.
My object, in writing the book, has been to make it useful to those who wish to become acquainted with the broad outlines of what is at present known of the morphology of the Invertebrata ; though I have not avoided the incidental mention of facts connected with their physiology and their distribution. On the other hand, I have abstained from discussing questions of aetiology, not because I underestimate their importance, or am insensible to the interest of the great problem of Evolution; but because, to my mind, the growing tendency to mix up aetiological speculations with morphological generalizations will, if unchecked, throw Biology into confusion.
For the student, that which is essential is a knowledge of the facts of morphology; and he should recollect that generalizations are empty formulas, unless there is something in his personal experience which gives reality and substance to the terms of the propositions in which these generalizations are expressed.
[5] The dissection of a single representative of each of the principal divisions of the Invertebrata will give the student a more real acquaintance with their comparative anatomy than any amount of reading of this, or any other book. And I have endeavored to facilitate practical study by supplying a somewhat full description of individual forms, in the case of the more complicated types.
That the power of repeating a "Classification of Animals," with all the appropriate definitions, has anything to do with genuine knowledge is one of the commonest and most mischievous delusions of both students and their examiners.
The real business of the learner is to gain a true and vivid conception of the characteristics of what may be termed the natural orders of animals. The mode of arrangement, or classification, of these into larger groups is a matter of altogether secondary importance. As such, I have relegated this subject to a subordinate place in the last chapter; and I have thought it unnecessary, either to discuss the systems proposed by others, or to give reasons for passing over, in silence, my own former attempts in this direction.
Of the manifold imperfections in the execution of the task which I have set myself, few will be more sensible than I am; but I trust that the book, such as it is, may be of use to the beginner.
Those who desire to pursue the study of the Invertebrata further will do well to consult the excellent treatises of Von Siebold,1 Gegenbaur,2 and Claus;3 and the elabo[6]rate works of Milne-Edwards4 and Bronn,5 in which a very full bibliography will be met with. Dr. Rolleston’s valuable "Types of Animal Life," and the "Elementary Instruction in Practical Biology," by myself and Dr. Martin, will prove useful adjuncts to the appliances of the practical worker.
LONDON, June, 1877.
1 "'Lehrbuch der vergleiehenden Anatomie der wirbellosen Thiere," 1848. One of the best books on the subject ever written, and still indispensable. 2 'Grundzüugeü der verglelohenden Anntomie," 1870; and "Grumlriss der verglelchenden Anatomie," 1874. 3 "Grundzuge der Zoologie." 3tte Auflage, 1879 4
'Leçons sur la Physiologie et l’Anatomie comparee de l’homme et des Animaux." Tomes i.-xii. (incomplete). 5 " Die Klassen und Ordnungen des Thierreichs." Bde. i.-vi. (incompIete).
[7]
| Preface | 8 | |
| Introduction to the General Principles of Biology | 9 | |
| Chap. I | The Distinctive Characters of Animals | 44 |
| II | The Protozoa | 73 |
| III | The Porifera and the Cœlenterata | 102 |
| IV | The Turbellaria, the Rotifera, the Trematoda, and the Cestoidea | 157 |
| V | The Hirudinea, the Oligochæta, the Polychæta, and Gephyra | 189 |
| VI | The Arthropoda | 219 |
| VII | The Air-Breathing Arthropoda | 320 |
| VIII | The Polyzoa, the Brachipoda, and the Mollusca | 389 |
| IX | The Echinodermata | 466 |
| X | The Tunicata or Ascidioda | 510 |
| XI | The Peripatidea, the Myzostomata, the Enteropneusta, the Chaætognatha, the Nematoidea, the Physemaria, the Acanthocephala, and the Dicyemida | 534 |
| XII | The Taxonomy of Invertebrated Animals | 561 |
| Index | 589 |
[9]
I.–The General Principles of Biology.
The biological sciences are those which deal with the phenomena manifested by living matter; and though it is customary and convenient to group apart such of these phenomena as are termed mental, and such of them as are exhibited by men in society, under the heads of Psychology and Sociology, yet it must be allowed that no natural boundary separates the subject-matter of the latter sciences from that of Biology. Psychology is inseparably linked with Physiology; and the phases of social life exhibited by animals other than man, which ‘sometimes curiously foreshadow human policy, fall strictly within the province of the biologist.
On the other hand, the biological sciences are sharply marked off from the abiological, or those which treat of the phenomena manifested by not-living matter, in so far as the properties of living matter distinguish it absolutely from all other kinds of things, and as the present state of knowledge furnishes us with no link between the living and the not-living.
These distinctive properties of living matter are–
1. Its chemical composition–containing, as it invariably does, one or more forms of a complex compound of carbon, hydrogen, oxygen, and nitrogen, the so-called protein (which has never yet been obtained except as a product of living bodies) united with a large proportion of water, and forming [10] the chief constituent of a substance which, in its primary unmodified state, is known as protoplasm.
2. Its universal disintegration and waste, by oxidation; and its concomitant reintegration by the intussusception of new matter.
A process of waste resulting from the decomposition of the molecules of the protoplasm, in virtue of which they break up into more highly-oxidated products, which cease to form any part of the living body, is a constant concomitant of life. There is reason to believe that carbonic acid is always one of these waste products, while the others contain the remainder of the carbon, the nitrogen, the hydrogen, and the other elements which may enter into the composition of the protoplasm.
The new matter taken in to make good this constant loss is either a ready-formed protoplasmic material, supplied by some other living being, or it consists of the elements of protoplasm, united together in simpler combinations, which consequently have to be built up into protoplasm by the agency of the living matter itself. In either case, the addition of molecules to those which already existed takes place, not at the surface of the living mass, but by interposition between the existing molecules of the latter. If the processes of disintegration and of reconstruction which characterize life balance one another, the size of the mass of living matter remains stationary, while, if the reconstructive process is the more rapid, the living body grows. But the increase of size which constitutes growth is the result of a process of molecular intussusception, and therefore differs altogether from the process of growth by accretion, which may be observed in crystals and is effected purely by the external addition of new matter–so that, in the well-known aphorism of Linnaeus,1 the word "grow," as applied to stones, signifies a totally different process from what is called "growth" in plants and animals.
3. Its tendency to undergo cyclical changes.
In the ordinary course of Nature, all living matter proceeds from preexisting living matter, a portion of the latter being detached and acquiring an independent existence. The new form takes on the characters of that from which it arose; exhibits the same power of propagating itself by means of an offshoot; and, sooner or later, like its predecessor, ceases to [11] live and is resolved into more highly-oxidated compounds of its elements.
Thus an individual living body is not only constantly changing its substance, but its size and form are undergoing continual modifications, the end of which is the death and decay of that individual; the continuation of the kind being secured by the detachment of portions which tend to run through the same cycle of forms as the parent. No forms of matter which are either not living, or have not been derived from living matter, exhibit these three properties, nor any approach to the remarkable phenomena defined under the second and third heads. But, in addition to these distinctive characters, living matter has some other peculiarities, the chief of which are the dependence of all its activities upon moisture and upon heat, within a limited range of temperature, together with the fact that it usually possesses a certain structure, or organization.
As has been said, a large proportion of water enters into the composition of all living matter; a certain amount of drying arrests vital activity, and the complete abstraction of this water is absolutely incompatible with either actual or potential life. But many of the simpler forms of life may undergo desiccation to such an extent as to arrest their vital manifestations and convert them into the semblance of not-living matter, and yet remain potentially alive; that is to say, on being duly moistened they return to life again. And this revivification may take place after months, or even years, of arrested life.
The properties of living matter are intimately related to temperature. Not only does exposure to heat sufficient to decompose protein matter destroy life, by demolishing the molecular structure upon which life depends; but all vital activity, all phenomena of nutritive growth, movement, and reproduction, are possible only between certain limits of temperature. As the temperature approaches these limits the manifestations of life vanish, though they may be recovered by return to the normal conditions; but, if it pass far beyond these limits, death takes place.
This much is clear; but it is not easy to say exactly what the limits of temperature are, as they appear to vary in part with the kind of living matter, and in part with the conditions of moisture which obtain along with the temperature. The conditions of life are so complex in the higher organisms at the experimental investigation of this question can be [12] satisfactorily attempted only in the lowest and simplest forms. It appears that, in the dry state, these are able to bear far greater extremes both of heat and cold than in the moist condition. Thus Pasteur found that the spores of fungi, when dry, could be exposed without destruction to a temperature of 120°-125° C. (248°-257° Fahr.), while the same spores, when moist, were all killed by exposure to 100° C. (212° Fahr.). On the other hand, Cagniard de la Tour found that dry yeast might be exposed to the extremely low temperature of solid carbonic acid (–60° C. or –76° Fahr.) without being killed. In the moist state he found that it might be frozen and cooled to –5° C. (23° Fahr.), but that it was killed by lower temperatures. However, it is very desirable that these experiments should be repeated, for Cohn's careful observations on Bacteria show that, though they fall into a state of torpidity, and, like yeast, lose all their powers of exciting fermentation at, or near, the freezing-point of water, they are not killed by exposure for five hours to a temperature below–10° C. (14° Fahr.), and, for some time, sinking to–18° C. (–0°.4 Fahr.). Specimens of Spirillum volutans, which had been cooled to this extent, began to move shout some little time after the ice containing them thawed. . . .
[15] The broad distinctions which, as a matter of fact, exist between every known form of living substance and every other component of the material world, justify the separation of the biological sciences from all others. But it must not be supposed that the differences between living and not-living matter are such as to bear out the assumption that the forces at work in the one are different from those which are to be met with in the other. Considered apart from the phenomena of consciousness, the phenomena of life are all dependent upon the working of the same physical and chemical forces as those which are active in the rest of the world. It may be convenient to use the terms "vitality" and "vital force" to denote the causes of certain great groups of natural operations, as we employ the names of "electricity" and "electrical force" to denote others; but it ceases to be proper to do so, if such a name implies the absurd assumption that either "electricity" or "vitality" is an entity playing the part of an efficient cause of electrical or vital phenomena. A mass of living protoplasm is simply a molecular machine of great complexity, the total results of the working of which, or its vital phenomena, depend, on the one hand, upon its construction, and, on the other, upon the energy supplied to it; and to speak of "vitality" as anything but the name of a series of operations is as if one should talk of the horologity" of a clock.
Living matter, or protoplasm and the products of its metamorphosis, may be regarded under four aspects:
(1.) It has a certain external and internal form, the latter being more usually called structure;
(2.) It occupies a certain position in space and in time;
(3.) It is the subject of the operation of certain forces, in virtue of which it undergoes internal changes, modifies external objects, and is modified by them; and–
(4.) Its form, place, and powers, are the effects of certain causes.
In correspondence with these four aspects of its subject, Biology is divisible into four chief subdivisions–I. MORPHOLOGY; II. DISTRIBUTION; III. PHYSIOLOGY; IV. AETIOLOGY.
....
Histology, which embodies the results of this analysis, shows that every tissue of a plant is composed of more or less modified structural elements, each of which is termed a cell; which cell, in its simplest condition, is merely a spheroidal mass of protoplasm, surrounded by a coat or sac–the cell-wall–which contains cellulose. In the various tissues, these cells may undergo innumerable modifications of form–the protoplasm may become differentiated into a nucleus with its nucleolus, a primordial utricle, and a cavity filled with a watery fluid, and the cell-wall may be variously altered in composition or in structure, or may coalesce with others. But, however extensive these changes may be, the fact that the tissues are made up of morphologically distinct units–the cells–remains patent. And, if any doubt could exist on the subject, it would be removed by the study of development, which proves that every plant commences its existence as a simple cell, identical in its fundamental characters with the less modified of those cells of which the whole body is composed.
But it is not necessary to the morphological unit of the plant that it should be always provided with a cell-wall. Certain plants, such as Protococcus, spend longer or shorter periods of their existence in the condition of a mere spheroid of protoplasm, devoid of any cellulose wall, while, at other times, the protoplasmic body becomes inclosed within a cell-wall, fabricated by its superficial layer.
Therefore, just as the nucleus, the primordial utricle, and the central fluid, are no essential constituents of the morphological unit of the plant, but represent results of its metamorphosis, so the cell-wall is equally unessential; and either the term "cell" must acquire a merely technical significance [18] as the equivalent of morphological unit, or some new term must be invented to describe the latter. On the whole, it is probably least inconvenient to modify the sense of the word "cell."
The histological analysis of animal tissues has led to similar results, and to difficulties of terminology of precisely the same character. In the higher animals, however, the modifications which the cells undergo are so extensive that the fact that the tissues are, as in plants, resolvable into an aggregation of morphological units, could never have been established without the aid of the study of development, which proves that the animal, no less than the plant, commences its existence as a simple cell, fundamentally identical with the less modified cells which are found in the tissues of the adult.
Though the nucleus is very constant among animal cells, it is not universally present; and, among the lowest forms of animal life, the protoplasmic mass which represents the morphological unit may be, as in the lowest plants, devoid of a nucleus. In the animal the cell-wall never has the character of a shut sac containing cellulose; and it is not a little difficult, in many cases, to say how much of the so-called "cell-wall" of the animal cell answers to the "primordial utricle" and how much to the proper "cellulose cell-wall" of the vegetable cell. But it is certain that in the animal, as in the plant, neither cell-wall nor nucleus is an essential constituent of the cell, inasmuch as bodies which are unquestionably the equivalents of cells–true morphological units–may be mere masses of protoplasm, devoid alike of cell-wall and nucleus
For the whole living world, then, it results: that the morphological unit–the primary and fundamental form of life–is merely an individual mass of protoplasm, in which no further structure is discernible; that independent living forms may present but little advance on this structure, and that all the higher forms of life are aggregates of such morphological units or cells variously modified.
. . . [19] In the course of its development every cell proceeds, from a condition in which it closely resembles every other cell, through a series of stages of gradually-increasing divergence, until it reaches that condition in which it presents the characteristic features of the elements of a special tissue. The development of the cell is, therefore, a gradual progress from the general to the special state.
The like holds good of the development of the body as a whole. However complicated one of the higher animals or plants may be, it begins its separate existence under the form of a nucleated cell. This, by division, becomes converted into an aggregate of nucleated cells–the parts of this aggregate, following different laws of growth and multiplication, give rise to the rudiments of the organs; and the parts of these rudiments again take on those modes of growth, multiplication, and metamorphosis, which are needful to convert the rudiment into the perfect structure.
The development of the organism as a whole, therefore, repeats in principle the development of the cell. It is a progress from a general to a special form, resulting from the gradual differentiation of the primitively similar morphological units of which the body is composed.
Moreover, when the stages of development of two animals are compared, the number of these stages which are similar to one another is, as a general rule, proportional to the closeness of the resemblance of the adult forms, whence it follows that the more closely any two animals are allied in adult structure, the later are their embryonic conditions distinguishable. And this general rule holds for plants no less than for animals. . . .
[235]
The Trilobita.–These ancient Arthropods, which have been extinct since the latter part of the Palæozoic epoch, occur in the fossil state in great numbers, and in conditions very favorable for their preservation; but, up to this time, no certain indications of the existence of appendages, nor even of any hard, eternal body-wall, have been discovered, though [236] a shield-shaped labrum, which lies in front of the mouth, has been preserved in some specimens. The body consists of a cephalic shield (Fig. 57, A); of a variable number of movably-articulated thoracic somites (Fig. 57, B); and of a pygidium, composed of a variable number of the somites which succeed the thorax, united together (Fig. 57, C).
Each thoracic somite presents a median portion, convex from side to side, termed the axis or tergum, and two flattened lateral portions, the pleura. The former overlap one another largely when the body is extended, the latter when it is flexed, and the freedom of motion permitted by this arrangement is so great that many Trilobites were able to roll themselves up like wood-lice, and are found fossilized in that condition. At the lateral edge of each pleuron, the cuticular substance of which it is composed folds inward, and can be traced on the ventral or sternal side for some distance. But in the middle of the ventral region no indication of a sternum is discoverable. It may, therefore, be concluded that the sternal region of the somite was of a soft and perishable nature; and that the thoracic somite of a Trilobite resembled one of the abdominal somites of a crab in this and in some other respects.
The glabellum (Fig. 57, 4), or central raised ridge of the cephalic shield, is a continuation of the thoracic axis, the location of its sides perhaps referring to the number of primitive somites it represents. The limb, or lateral area on either side, answers to a thoracic pleuron; its thickened margin (Fig. 57, 1) is produced into two longer or shorter posterior angles (g); inferiorly, the marginal band is reflected inward for a short distance, as the subfrontal fold, the remaining sternal area being incomplete. A median movable plate answers to the labrum of Apus and Limulus. On the occipital or lateral margin of the limb a suture (Fig. 57, 5) commences, and, passing between the eye and the glabellum, meets that of the opposite side either in front of the latter, or on the margin of the limb, or on the subfrontal fold, and is connected with the labral suture by one or two sutures. The limb is thus divided into two parts–one fixed (the fixed gena, Fig. 57, a), attached to the glabellum; the other separated (the movable gena, Fig. 57, b), on which the eye is placed. The eyes are absent in some genera. In others they occur as isolated ocelli; or in groups, their interspaces being occupied by the common integument; or they may resemble the compound eyes of other Arthropods.
[237] M. Barrande1 has succeeded in tracing out the development of some species of Trilobites. He finds that the smallest, and therefore the youngest, forms are discoidal bodies, without any clear evidence of segmentation. The division into somites takes place by degrees, the number increasing up to the adult condition. It is possible that still younger conditions may have escaped fossilization, but the analogy of Limulus suggests that these small discoidal forms really represent the condition in which the Trilobite left the egg.
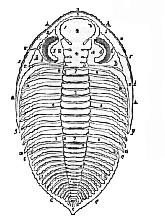
A, head; 1, marginal band; 2. marginal groove, internal to the band; 3. occipital segment; 4, glabellum; 5, great suture; 6, eyes; a, fixed gena; b. separable gena; g. genal angle;
B, thorax; 7, axis or tergum; 8, pleuron;
C, pygidium; 9, tergal; 10, pleural portions of the pygidium.
The Merostomata.2–The only existing representative of this division of the Crustacea is the genus Limulus (the King Crabs or Horseshoe Crabs), the various species of which are [238] found in America and in the Moluccas. They are usually classed as a distinct order of the Crustacea, termed Xiphosura or Pœcilopoda.
The body of Limulus (Fig. 58) is naturally divided into three parts, which are movably articulated together. The most anterior is a shield-shaped portion, curiously similar in form to the head of a Trilobite. Its convex dorsal surface is similarly divided into a median and two lateral regions; its edges are thickened, and its posterior and external angles are produced backward. At the anterior end of the median region two simple eyes are situated, and at its sides are two large compound eyes. The sternal surface presents, anteriorly, a flattened subfrontal area, behind which it is deeply excavated, so that the labrum and the appendages are hidden in a deep cavity formed by its shelving walls. The middle division of the body of Limulus exhibits markings which indicate that it is composed of, at fewest, six coalesced somites; its margins are spinose, and its excavated sternal face lodges the appendages of this region.
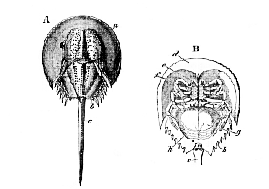
B. L. rotundicauda (ventral view) (after Milne-Edwards): a, anterior, b, middle division of the body, c, telson, d, subfrontal area; e, antennules; f, antennæ; g, operculum; h, branchiferous appendages.
The terminal division is a long, pointed, and laterally serrated spine, which is termed the telson.
[229] The mouth is placed in the centre of the sternal surface of the anterior division; the anus opens on the same surface, at the junction between the middle division and the telson. A movable, escutcheon-shaped labrum projects backward in the middle line, immediately behind the subfrontal area (d); and on each side of it is a three-jointed appendage, the second joint of which is prolonged in such a manner as to form with the third a pincer or chela. The attachment of this appendage is completely in front of the labrum, which separates it from the mouth.
In each of the next five pairs of appendages, the basal joint is enlarged; and, in the anterior four, its inner edge is beset with numerous movable spines. The attachment of the basal joint of the foremost of these appendages (the second of the whole series) is in front of the mouth; but its prolonged, spinose, posterior and internal angle may be made to project a little into the oral cavity. The basal joints of the following three appendages are articulated at the sides of the mouth, and the inner angle of each is provided with a spinose process which projects into the oral cavity. The second, third, fourth, and fifth appendages in the females are chelate; in the males of most species the second, and sometimes the third, are not chelate. The large basal joint of the sixth appendage is almost devoid of spines, and bears a curved, spatulate process, which is directed backward between the anterior and middle divisions of the body. The fifth joint of this limb carries four oval lamellæ. The appendages which form the seventh pair, very unlike the rest, are short, stout, and single-jointed.
The eighth pair of appendages, again, are of a totally different character from those which precede them. They are united in the middle line into a single broad plate, which forms a sort of cover, or operculum, over the succeeding appendages, when the animal is viewed from the sternal side. On the dorsal face of this plate are seated the two apertures of the reproductive organs....
1 "Systéme Silurien du centre de Bohème," tome 1. Trilobites. 1852. 2 H. Woodward, "A Monograph of the British Fossil Crustacea belonging to the Order Merostomata," 1866.
[440] The Pulmonata.–These are odontophorous Mollusks which breathe air directly by means of a respiratory surface furnished by the wall of the pallial cavity.
In some, such as the Peroniadæ (Fig. 123) and Veronicellidæ, the body of the slug-like animal is very nearly symmetrical; the anus and the lung-sac being situated close together at the posterior extremity of the body. The mantle is large, and extends over the whole hæmal or dorsal surface. In all the other Pulmonata, the pulmonary and the anal apertures lie on the right side of the body, and the mantle is provided with at least the rudiments of a shell. The pallial region is sometimes very small in proportion to the rest of the body, and then forms a flattened disk, as in the common Slug; while, in some Limacidæ and Testacellidæ, and in the [441] Janellidæ, the mantle is so much reduced that they are almost achlamydate. In the Snails, the mantle is large and is produced into an asymmetrically coiled visceral sac, in which the stomach, liver, and genital gland lie. The mantle-cavity lies on the fore-part of the sac, and the anus opens on its margin. Thus, in all the ordinary Pulmonata, the termination of the intestine is twisted from its normal position at the hinder end, forward to the right dorsal, or hæmal, aspect of the body.
When the pulmonary sac is posterior, and the pallial region small, the ventricle of the heart is anterior, and the auricle posterior, and the animal may be said to be opisthopulmonate. On the other hand, when the pallial region is large, and gives rise to a visceral sac, with the concomitant forward position of the pulmonary chamber, the auricle is inclined more or less forward and to the right side, and the apex of the ventricle backward and to the left side. The animal is thus more or less prosopulmonate.
The mouth is commonly provided with a horny upper jaw, as well as with a well-developed odontophore. Large salivary glands are usually present.
The heart consists of a single auricle and a single ventricle. The aortic trunk, which proceeds from the apex of the latter, divides into many branches, but the venous channels are altogether lacunar. A renal organ lies close to the pulmonary sac in the course of the current of the returning blood.
There are usually two simple eyes, often lodged in the summits of retractile tentacula.
The Pulmonata are hermaphrodite. The generative gland is an ovotestis, and is composed of branched tubuli, from the cellular contents of which both ova and spermatozoa are developed (Fig. 123, III.).
A narrow common duct leads from the ovotestis, and, soon dilating, receives the viscid secretion of a large albumen-gland. The much wider portion of the common duct beyond the attachment of this gland is incompletely divided by longitudinal infoldings into a sacculated, wider, and a straight, narrower, division. The former conveys the ova, and the latter the spermatozoa. At the end of this part of the apparatus, the wider portion, which represents the oviduct, passes into the vagina, which opens at the female genital aperture, while the narrower portion of the common duct is continued into a separate, narrow, vas deferens, the end of which opens into a long invagination of the integument–the [442] penis. In Peronia, the vas deferens and the oviduct open together by the genital aperture, and, as in some Branchiogasteropods, a groove, along which the seminal fluid is conducted, leads to the outer opening of the eversible penis (Fig. 123, I., II.).
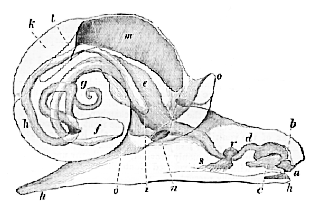
Fig. 122.–Diagram exhibiting the disposition of the intestine, nervous system, etc., in a common Snail (Helix).–a, mouth; b. tooth; e, odontophore; d, gullet; e, its dilatation into a sort of crop; f, stomach; g, coiled termination of the visceral mass; the latter is also close to the commencement of the intestine, which will be seen to lie on the neural side of the oesophagus; h, rectum; i, anus; k, renal sac; l, heart; m, lung, or modified pallial chamber; n, its external aperture; o, thick edge of the mantle united with the sides of the body; p, foot; r, s, cerebral, pedal, and parieto-splanchnic ganglia aggregated round the gullet.
In connection with the female genital aperture, there is always a spermatheca, or sac (which is sessile in the Slugs, but in the Snails is placed at the extremity of a long duct), for the reception of the semen of the other individual when copulation takes place.
The Helicidæ alone possess, in addition, the so-called sac of the dart, a short muscular bag, in which pointed chitinous or calcified bodies–the spicula amoris–are formed; and certain glandular cæca, generally arranged in two digitate bundles, termed mucous glands, which give rise to a milky secretion. Sometimes prostatic glands are developed on the [443] vas deferens, which may be dilated in part of its course into a vesticula seminalis.
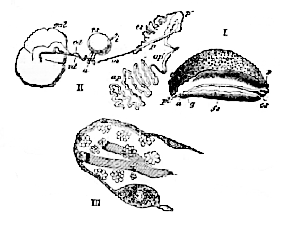
I. Peronia verruculata,–a, anus; pl. pulmonary aperture; g, genital aperture; fs, seminal groove; p, opening for the penis.
II. Generative organs of the same animal, the ovotestis being omitted.–gal, gland which furnishes a glairy secretion; od, oviduct; vd, vas deferens; i, intestine; a, anus; rs, receptaculum seminis; p. aperture of the penis; p', penis; cs, seminal duct; ap, glandular appendage; m, retractor muscle of the penis. (After Keferstein.)
III. Blind end of a follicle of the ovotestis of Helix pomatia. At the apex the spermatozoa are seen in different stages of development, the fully-formed spermatozoa floating in bundles in the cavity of the follicle. Lower down, ova are developing in the walls of the follicle. (After Keferstein and Ehiers.)
The ova are impregnated high up in the oviduct, and are invested by a relatively very large mass of albumen and inclosed within a thick, sometimes calcified, chorion. The mass inclosed by the latter may be a tenth of an inch or more in diameter, while the proper ovum may have not more than a twelfth of that size.
There is no trustworthy evidence of the existence of the opisthobranchiate Gasteropods before the epoch of the Trias, but it is to be remembered that the great majority of these animals have no shells. Of the rest of the preceding groups of Odontophora, representatives are known as far back as the middle of the Palæozoic epoch, while Pteropoda, Hetero[444]poda, and Prosobranchiata, occur in the Silurian formations. Among the Prosobranchiata, the Patellidæ and the Aspidobrachia are the characteristic forms of the oldest formations, the Ctenobranchia appearing later, and acquiring their present relative abundance only in the later secondary and the tertiary epochs.
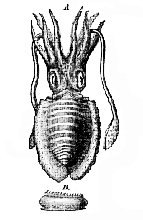
The Cephalopoda.– – The bilateral symmetry which is so obvious in the Polyplacophora and the Scaphopoda is but little disturbed in this group of the Odontophora. The mouth and the anus are situated in the median plane, which divides the body into corresponding halves; while the branchiæ, two or four in number, are disposed symmetrically on each side of this plane, as are the brachial prolongations of the margins of the foot. The hæmal face of the body, however, is not flat, as in the mollusks which have just been mentioned, but is elongated perpendicularly to the neural face, so as to form a sort of sac, invested by the mantle. On the pos[443]terior, or anal, face of the sac, the mantle incloses a large pallial cavity, in which the branchiæ are protected. On the anterior aspect of the sac, on the contrary, the mantle may have no free edge, or, at most, forms a comparatively small flap.1
The integument is provided with chromatophores, which are sacs with elastic walls, full of pigment, and provided with radiating muscles, by which they may be drawn out to a size many times greater than that which they possess in their contracted state. In their dilated condition, the color proper to the contained pigment becomes plainly visible, while in their contracted state they appear as mere dark specks. It is to the successive expansion and contraction of these chromatophores that the Cephalopoda owe the peculiar play of "shot" colors, which pass like blushes over their surface in the living state. These blushes of color are especially well displayed by young Cephalopods just freed from the egg.
But that which particularly distinguishes the Cephalopod is the form and disposition of the foot. The margins of this organ are, in fact, produced into eight or more processes, termed arms, or brachia ; and its antero-lateral portions have grown over and united in front of the mouth, which thus comes, apparently, to be placed in the centre of the pedal disk. Moreover, two muscular lobes which correspond with the epipodia of the Pteropods and Brachiogasteropods, developed from the sides of the foot, unite posteriorly, and, folding over, give rise to a more or less completely tubular organ, the funnel, or infundibulum. . . .
[582] The Serial Relations of the Invertebrata Compared with the Results of Embryology.–The conception of the unity of organization of the Invertebrata thus reached, so far as it is based upon the comparison of adult structures, is purely ideal; and the study of the development of individual animals is alone competent to decide the question whether this ideal unity has a foundation in objective fact. But the history of the development of animals appertaining to every group of the Invertebrata which has been given, bears out the statement which is made in the Introduction, that the ideal unity has such a foundation in fact; inasmuch as all these animals commence their existence under the same form–that, namely, of a simple protoplasmic body, the ovum or germ.
In the Introduction I have said that, "among the lowest forms of animal life, the protoplasmic mass which represents the morphological unit may be, as in the lowest plants, devoid of a nucleus" (p. 18). However, as I have remarked at the commencement of this chapter, until the search for the nucleus has been instituted afresh, with the help of such methods as have recently proved its existence in the Foraminifera, I think it will be wise to entertain a doubt whether any of the Monera are really devoid of this amount of structural differ[583]entiation; and the tendency of recent investigations appears to render it very questionable whether the nucleus of the ovum ever really disappears, whatever may be the modifications undergone by the germinal vesicle and its contents. I shall, therefore, assume provisionally, that the primary form of every animal is a nucleated protoplasmic body, cytode, or cell, in the most general acceptation of the latter term.
Whether the primary cytode possesses a nucleus or not, the important fact remains that, in its earliest condition, every invertebrated animal, if it were competent to lead an independent existence, would be classed among the Protozoa.
The first change which takes place in the development of the embryo from the primitive cytode, or impregnated ovum, in all the Metazoa, is its division; and the simplest form of division results in the formation of a spheroidal or discoidal mass of equal, or subequal, derivative cytodes, the blastomeres. Next, the morula, talus formed, generally acquires a central cavity the blastocoele, and becomes a hollow vesicle, the blastosphere, the wall of which, composed of a single layer of blastomeres, is the blastoderm.
The blastomeres of the blastoderm next undergo differentiation into two kinds, distinguished by their internal activities, if not by their outward form. Of these the one set constitute the epiblast, the others the hypoblast. The further changes of the embryo are the consequences of the tendencies toward further modification resident in the epiblastic and hypoblastic blastomeres respectively. Each of these is, as it were, a germ, whence certain parts of the adult organism will be evolved.
Every series of the Invertebrata has now yielded a number of examples of the further modification of the blastosphere by the process of invagination, or emboly, the result of which is that the hypoblast becomes more or less completely inclosed within the epiblast. The invagination is accompanied by the diminution, or even abolition, of the blastocoele, and the formation of a cavity inclosed within the hypoblast, which is the archenteron, or primitive alimentary cavity. The opening left by the approximated edges of the epiblast, when the process of invagination is completed, and by which the archenteron communicates with the exterior, is the blastopore. In this state the embryo is a gastrula.
It very commonly happens that the process of development is modified by an inequality in the size of the blastomeres; which inequality may be manifest from the bisection [584] of the ovum, or may appear later. In this case, it usually happens that the smaller and more rapidly-dividing blastomeres belong to the epiblast, and the larger and more slowly dividing to the hypoblast. Moreover, no blastocoele may arise, and the process of inclusion of the hypoblast within the epiblast may have the appearance of the growth of the latter over the former, or what is termed epiboly; while the archenteron may not be formed within the hypoblast till very late.
When, in cases of epiboly, the blastoderm is small in relation to the vitellus, the epiblast and hypoblast, at their first appearance, necessarily adapt themselves to the surface of the yelk; and thus the gastrula, instead of having the form of a deep cup, becomes more or less flattened and discoidal.
I am inclined to believe that all the various processes by which the gastrula or its equivalent are produced, are reducible to epiboly and emboly. Even when the epiblast and the hypoblast appear to be formed by delamination, or the splitting into two layers of cells of a primitively single-layered blastoderm, there seems little doubt that what happens is either the very early inclusion of the hypoblastic blastomeres within those which give rise to the epiblast, or a very late and inconspicuous ingrowth, or invagination, of the hypoblastic region of the blastoderm.
If we employ the term gastrula in the broad sense defined above, it may be truly said that every metazoon passes through the gastrula stage in the course of its development. The question whether the mode of development of the gastrula by emboly is primitive, and that by epiboly secondary; or whether epiboly is primary and emboly secondary; or whether the two processes have originated independently, is of secondary importance, and belongs to the debatable ground of phylogeny.1
. ... [586] It is very important to remark that these morphological generaliztions, so far as they are correctly made, are simple statements of fact, and have nothing to do with any speculations respecting the manner in which the invertebrated ani[587]mals with which we are acquainted have come into existence. They will remain true, so far as they are true at all, even if it should be proved that every animal species has come into existence by itself and without reference to any other. On the other hand, if there are independent grounds for a belief in evolution, the facts of morphology not ony present no difficulty in the way of the hypothesis of the evolution of the Invertebrata from a common origin, but readily adapt themselves to it.
Hence the numerous phylogenic hypotheses which have of late come into existence, and of which it may be said that all are valuable, so far as they suggest new lines of investigation, and that few have nay other significance. I do not desire to add to the number of these hypotheses; and I will only venture to remark that, in the absence of any adequate palaeontological history of the Invertebrata, any attempt to construct their Phylogeny must be mere speculation.
But the oldest portion of the geological record does not furnish a single example of a fossil which we have any reasonable grounds for supposing to be the representative of the earliest form of any one of the series of invertebrated animals; nor any means of checking our imaginations of what may have been, by evidence of what has been, the early history of invertebrates on the globe.
Already indications are not wanting that the vast multitude of fossil Arthropods, Mollusks, Echinoderms, and Zoophytes, now known, will yield satisfactory evidence of the filiation of successive forms, when the investigations of palaeontologists are not merely actuated by the desire to discover geological time-marks and to multiply species, but are guided by that pereception of the importance of morphological facts which can only be conferred by a large and thorough acquaintance with anatomy and embryology. But, under this aspect, the palaeontology of the Invertebrata has yet to be created.
1 Compare Haeckel, "Studien zur Gastræa-Theorie," in his "Biologische Studien," 1877.
|
THE
HUXLEY
FILE

|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||
{kind=link}